Содержание
БОТАНИЧЕСКИЕ ИСТОРИИ | Наука и жизнь
В литературе накопилось достаточное количество недоразумений, связанных с ботаникой, многие из которых имеют историю в несколько столетий.
ИСТОРИЯ ПЕРВАЯ, ИЛИ ЗАГАДОЧНАЯ ЦИКУТА
Наука и жизнь // Иллюстрации
Так выглядит особенно ядовитое растение — цикута; листья у нее, как у петрушки, а красивые цветки собраны в зонтики.
Наука и жизнь // Иллюстрации
Цветки аквилегии, или водосбора, орлика. Раскрытый цветок напоминает мирного голубя, бутон — хищную, нахохлившуюся птицу.
С мая по июнь почти по всей Европе на полях, пустырях, обочинах дорог, а также в посевах зерновых цветет ромашка аптечная.
В июне-июле на цветоносах, очень похожих на листья, появляются соцветия аира в виде початков с мелкими, неприметными зеленоватыми цветками. Растение можно найти по берегам прудов, в заболоченных канавах, болотистых местах.
Купырь лесной — обычное растение лесов и лугов Европы, Северной и Средней Азии, где может образовывать большие заросли. Цветет в июне-июле.
Цветет в июне-июле.
‹
›
Карл Линней, будучи гениальным ботаником и создателем системы растительного
и животного мира, в детстве, кроме ботаники, ничем не интересовался, за что
его чуть не отдали в учение к сапожнику. Позднее он вынужден был освоить некоторые
другие области знаний, весьма далекие от ботаники, например пробирное дело,
минералогию, и даже изобрел стоградусную шкалу термометра, которая сейчас известна
как шкала Цельсия. Что касается географии, знания его оставались весьма минимальными;
именно из-за этого в названиях растений возникло немало недоразумений. Так,
уже несколько столетий врачи и ботаники разбираются, чем отравили Сократа, поскольку
Линней назвал цикутой растение, которое в Греции не встречается. Судя по названиям
некоторых других растений, Линней был уверен, что Сибирь расположена сразу за
границей Польши. Среди числящихся, по Линнею, сибирскими очень многие растения
там и не встречаются.
ИСТОРИЯ ВТОРАЯ,
ИЛИ ВОДОСБОР, КОТОРЫЙ
НЕ СОБИРАЕТ ВОДУ
В декоративном садоводстве пользуется известностью красивое растение — водосбор (его научное название — аквилегия). Происхождение «имени» пытаются объяснить тем, что растение якобы удерживает воду в цветках или на листьях. На самом деле водосбор намокает под дождем не более других растений — известная многим манжетка удерживает воду гораздо лучше. Да и цветки у водосборов, встречающихся в отечественной флоре, устроены так, что вода не попадает на тычинки и пестики. В Америке есть водосборы с цветками, повернутыми вверх, но и они перед дождем поникают.
Откуда же взялось это странное название?
До середины XVIII века в большинстве стран, в том числе и в России, это растение называли либо голубками, либо орликами. «Птиц» нетрудно заметить, если рассмотреть цветок или крупный бутон. Чашелистики как бы образуют крылья, а лепесток, вытянутый в нектарник, — голову и шею. В зависимости от того, что мы рассматриваем — цветок или бутон, «птицы» выглядят по-разному: бутоны больше напоминают хищную нахохлившуюся птицу, а раскрытые цветки — мирного голубя.
Название «водосбор» родилось в середине XVIII века, вместе с появлением научной номенклатуры — официальных названий растений. В русской науке в это время господствовали немецкие ученые, многие из которых внесли в нее большой вклад. Но водосбору не повезло: его, по всей видимости, описывал человек, не знающий ни цветка, ни его русского названия, ни латинского языка. У немцев цветок носит название, производное от латинского слова «Аkelei» (акелей). Латинское название тоже «птичье»: аquila — по-латыни орел, соответственно Aquilegia — тот же русский орлик. Переводчик же усмотрел в этом слове сложное сочетание из слов аqua — вода и lego — собираю, так «вырос» на русской земле немецкий водосбор. А в народе как звали его орликом, так и зовут. Кстати, в словаре Даля он упоминается под обоими названиями.
ИСТОРИЯ ТРЕТЬЯ, ИЛИ ОТКУДА ВЗЯЛИСЬ РОМАШКИ
Ромашка — одно из немногих растений, известных любому говорящему на русском языке. Назвали этот цветок ромашкой сравнительно недавно. Раньше растения с белыми лепестками и желтой серединкой называли пупавками, поскольку в центре соцветия возвышается желтый «пупок». Название «ромашка» связано с проникновением на Русь средневековой медицинской литературы — травников и лечебников, в которых растения, сейчас называемые ромашкой, именовались «романом», «романовой травой», «романовым цветом».
Раньше растения с белыми лепестками и желтой серединкой называли пупавками, поскольку в центре соцветия возвышается желтый «пупок». Название «ромашка» связано с проникновением на Русь средневековой медицинской литературы — травников и лечебников, в которых растения, сейчас называемые ромашкой, именовались «романом», «романовой травой», «романовым цветом».
От слова «роман», не без влияния имени собственного, образовалась уменьшительная форма «ромашка», впервые зафиксированная только в конце XVIII века в рецепте помещика и первого русского агронома А. Г. Болотова, рекомендовавшего от простуды «…один декокт (отвар). Одна часть буквицы, другая ромашки, третья шалфея».
ИСТОРИЯ ЧЕТВЕРТАЯ, ТОМАТНАЯ
Одна из самых забавных ошибок ботаников Старого Света — история с помидорами. Все знакомые им прежде растения семейства паслёновых, а их в Старом Свете меньше десятка, были в разной степени ядовиты.
Первый европейский ботаник, упомянувший в 1554 году помидор, итальянец Пьетро Андреа Маттиолли, из-за крупных плодов вначале отнес его к роду мандрагора, славящемуся своей ядовитостью. А поскольку в Европу попали томаты желтой окраски, они получили итальянское название «помо д’оро» — золотое яблоко. Позже помидор отнесли к роду паслён и назвали Solanum lycopersicum (паслён волчий персик). Да и современное его название не лучше — Lycopersicum aesculentum — волчий персик съедобный.
А поскольку в Европу попали томаты желтой окраски, они получили итальянское название «помо д’оро» — золотое яблоко. Позже помидор отнесли к роду паслён и назвали Solanum lycopersicum (паслён волчий персик). Да и современное его название не лучше — Lycopersicum aesculentum — волчий персик съедобный.
Красивые плоды томатов, свисающие нарядными гроздьями, вызвали интерес у любителей-цвето водов, и новые декоративные растения прочно обосновались в коллекциях ботанических садов и на клумбах. Французы назвали их «пом д’амур» — яблоко любви.
Кто из европейцев первым попробовал помидор и когда это произошло, неизвестно, но еще в XVIII веке это растение было малоупотребительно как пищевое. В 1780 году российский посол во Франции докладывал Екатерине II, что французские бродяги едят помидоры с клумб и вроде бы от этого не страдают. Более того, даже в Америке, на континенте, где перуанские и мексиканские индейцы уже давно выращивали томаты (название «туматль» вслед за мексиканцами впервые употребил в 1572 году итальянский ученый Гиландини), помидор до середины XIX века считался ядовитым. Причем настолько ядовитым, что в 1776 году, во время борьбы Америки за независимость, повар Джорджа Вашингтона попытался отравить его мясом, приготовленным с помидорами. Сам повар был настолько напуган содеянным, что перерезал себе горло в страхе перед наказанием, а Джордж Вашингтон, отведав томатного соуса, остался и жив, и доволен. Вот так ботаники, намудрившие с помидорами, надолго запугали европейцев их мнимой ядовитостью. Помидорный бум в мире начался только после Первой мировой войны — более чем через 350 лет после первого знакомства европейцев с помидорами.
Причем настолько ядовитым, что в 1776 году, во время борьбы Америки за независимость, повар Джорджа Вашингтона попытался отравить его мясом, приготовленным с помидорами. Сам повар был настолько напуган содеянным, что перерезал себе горло в страхе перед наказанием, а Джордж Вашингтон, отведав томатного соуса, остался и жив, и доволен. Вот так ботаники, намудрившие с помидорами, надолго запугали европейцев их мнимой ядовитостью. Помидорный бум в мире начался только после Первой мировой войны — более чем через 350 лет после первого знакомства европейцев с помидорами.
ИСТОРИЯ ПЯТАЯ. ОШИБКА ПОЭТА, ИЛИ СОРНЯК В МРАМОРНОМ БАССЕЙНЕ
Поэт и писатель А. К. Толстой, перу которого принадлежат общеизвестные «Колокольчики мои…», очень часто упоминал в своих произведениях различные растения. Но в стихотворении «Алеша Попович» он допустил любопытную ботаническую ошибку:
«Кто веслом так ловко
правит
Через аир и купырь?
Это тот Попович
славный,
Тот Алеша богатырь!»
Прекрасно рифмующийся со словом «богатырь», купырь, увы, растение хоть и влажных мест, но сухопутное, и «править» через него лодку невозможно.
Вторая и более интересная ошибка — упоминание аира. Легендарный Алеша Попович жил во времена Владимира Красное Солнышко, то есть в Х или начале XI века (Владимир умер в 1015 году), а аир появился на Руси значительно позже — только во время татарского нашествия. Татары считали его указателем чистой воды и бросали кусочки корневищ во все встречные водоемы. В словаре Даля указано его название — «татарская сабля». Интересно, что в Московской области аир встречается почти исключительно по реке Москве и ее крупнейшим притокам; вероятно, остальные леса были непреодолимы для татарских войск. Так что аира Алеша Попович увидеть не мог, не говоря уже о том, чтобы через него пробираться, для этого заросли должны быть большими.
Источником недоразумений аир был неоднократно. Нельзя сказать, чтобы аир не знали совсем. В Средние века его в довольно больших количествах ввозили из Турции в Европу в качестве пряности и лекарства, но образцы самого растения для ботаников долгое время были недоступны.
Только в 1557 году итальянский ботаник Маттиолли получил корневища аира, собранные в Турции на озере Никомедия, в качестве подарка от турецкого султана для императора Фердинанда. В 1576 году корневища прислали в Вену, на сей раз из Константинополя. Из этих двух садов аир начал распространяться по Западной Европе и уже в конце XVII века стал довольно обычным растением.
Рассказывают, что как-то в Вену заехал начинающий ботаник из Польши. Молодой человек с удивлением спросил директора сада, известнейшего ботаника К. Клузиуса, зачем он держит в роскошном бассейне такое обыкновенное растение, известное в любой польской деревне. Представьте себе удивление и возмущение знаменитого ботаника. Но студент был прав: аир, занесенный в Польшу татарами, рос там уже около 300 лет и успел стать вполне привычным растением. И до сегодняшнего дня его называют «татарак».
Годичное собрание 2021 г. | Общество физиологов растений
Всероссийская научная конференция с международным участием и школа для молодых ученых,
«Экспериментальная биология растений и биотехнология: история и взгляд в будущее»
посвященные 130-летию ИФР РАН и 100-летию со дня рождения чл. -корр. РАН Р.Г. Бутенко
-корр. РАН Р.Г. Бутенко
Москва, ИФР РАН
27 сентября — 1 октября 2021 г.
На конференции, посвященной 130-летнему юбилею ИФР РАН и 100-летию со дня рождения чл.-корр. РАН Р.Г. Бутенко будут представлены актуальные направления исследований в области современной биологии растений, а также последние достижения методологии исследований и их практического применения. В рамках мероприятия состоится школа молодых ученых. Вне научной программы конференции пройдет Годичное собрание ОФР.
ВАЖНО! ВНИМАНИЕ!
Уважаемые участники! В связи с ухудшившейся эпидемиологической обстановкой в Москве были приняты ряд карантинных ограничений. Конференция в ноябре не состоится и перенесена на 2021 год. Все тезисы и оргвзносы сохранены. Участники вправе отозвать свое участие и вернуть средства. Дата проведения конференции переносится на 27 сентября — 1 октября 2021 г., Москва.
Вопросы оргкомитету просим задавать через официальный адрес ifr2020@ofr. su
su
НАУЧНЫЕ НАПРАВЛЕНИЯ
- Регуляция экспрессии генома и трансдукция сигналов в процессах клеточной дифференцировке и онтогенеза растений
- Физиология, биохимия и экология фотосинтеза, дыхания и фиксации азота как теоретическая основа продукционного процесса
- Адаптация растений в связи с экологическими стрессами и глобальными биосферными явлениями
- Биология фототрофных и гетеротрофных клеток растений как основа развития инновационных биотехнологий
- Проблемы подготовки специалистов в области физиологии и биохимии растений
КЛЮЧЕВЫЕ ДАТЫ
| Первое информационное письмо | 20 февраля 2020 г. |
| Последний день приема заявок на участие и тезисов | 31 мая 2021 г. |
| Второе информационное письмо | 11 июня 2021 г. |
| Оплата участия | до 1 августа 2021 г. |
| Школа молодых ученых | |
| Смена очного участия на заочное или отмена с отзывом тезисов |
СТОИМОСТЬ УЧАСТИЯ
| Члены ОФР | НЕ члены ОФР | |
| Очное участие | 2 000 р | 2 500 р |
| Молодые ученые (не старше 35 лет) | 1 000 р | 1 500 р |
| Заочное участие или публикация вторых тезисов | 500 р | 500 р |
КОНТАКТЫ
| Место проведения | 127276, Москва, ул. Ботаническая 35 |
| Телефоны | +7 (499) 977-80-22 |
| [email protected] | |
| Адрес сайта | ofr.su/ifr2020 |
История горшков для растений — green-land.
 ru
ru
Все имеет свое начало
Цветочные горшки, кашпо и вазоны сегодня привычные предметы обихода и украшения в доме, саду и офисе, возникли в эпоху неолита (в 8-7 веках до Рождества Христова). Когда культуры, живущие за счет охоты и собирания плодов, стали оседлыми и перешли к земледелию и скотоводству, им понадобились большие горшки для хранения своих растений. В таких больших горшках, прототипах цветочных горшков, можно было удобно выращивать молодые ростки. Так как время от времени приходилось менять место жительства, то цветочные горшки в тот период использовались, прежде всего, для транспортировки растений.
Первые горшки
служили нашим предкам для хранения в них воды, продуктов питания и личных вещей. Сначала такие большие горшки делали из листьев, плетеной ивы, полых стволов деревьев или камня. Только открытие, что глина, лежащая вблизи огня, стала твердой и водонепроницаемой, привело к изготовлению из нее вазонов и цветочных горшков. Самые древние керамические горшки были обнаружены в 8 веке до Рождества Христова.
Одно открытие изменяет жизнь
С тех пор это открытие изменило жизнь человека. Со временем барельефы, мозаика и изразцы украсили здания внутри и снаружи, а цветы, обычно растущие под открытым небом, отлично ужились под крышей в цветочных горшках, вазонах и кашпо. Судьбоносная связь человека с этим первым, материалом, форму которому он придавал сам, отражается в мифах самых разных народов: почти у каждого из них есть предание о том, что бог-творец создал человека из глины и вдохнул в него жизнь. Так говорит и Библия в Книги Бытия 2.7: «И создал Господь Бог человека из праха земного, и вдунул в лице его дыхание жизни, и стал человек душою живою.»
В связи с этим прототипы горшков играли особую роль в ходе религиозных церемоний. Они украшались в соответствии со своим ритуальным назначением, и тогда оставался всего лишь шаг к использованию их в качестве декоративного украшения храмов, жилых и общественных зданий и садов.
Новое исследование проливает свет на «ужасную тайну» Дарвина, преследовавшую его до конца жизни
Автор фото, Getty Images
Подпись к фото,
В своей теории эволюции Чарльз Дарвин описал естественный процесс развития живой природы
Термин «ужасная тайна» Чарльза Дарвина — широко известен. Не секрет, что великий ученый так и не смог объяснить с точки зрения эволюции происхождение на Земле цветковых растений. Но лишь сейчас стало известно, что тайна цветов чуть не стоила Дарвину труда всей его жизни и угнетала его до последних дней.
Не секрет, что великий ученый так и не смог объяснить с точки зрения эволюции происхождение на Земле цветковых растений. Но лишь сейчас стало известно, что тайна цветов чуть не стоила Дарвину труда всей его жизни и угнетала его до последних дней.
Разбирая архивные документы, биолог-эволюционист из Лондонского университета королевы Марии, профессор Ричард Баггс обнаружил, что за несколько лет до смерти Дарвина у него появился весьма решительный оппонент — шотландский ботаник Уильям Каррутерс.
Каррутерс придерживался креационистской теории происхождения цветковых растений, полагая, что они возникли благодаря вмешательству свыше, и раструбил в прессе, что Дарвин не в состоянии дать по этому вопросу естественнонаучного объяснения.
Пробел в дарвиновской теории эволюции стал достоянием общественности и грозил подорвать позиции Дарвина в научном мире.
Тогда-то, считает Ричард Баггс, и родилось это словосочетание — the abominable mystery: ужасная или отвратительная тайна.
Что такое «ужасная тайна» Дарвина?
Впервые этот термин Чарльз Дарвин употребил в 1879 году в письме своему другу, исследователю и ботанику Джозефу Хукеру. В нем он писал, что стремительное по геологическим меркам развитие высших видов растений составляет ужасную тайну.
Речь шла о цветах и цветковых (или покрытосеменных) растениях, отличительная особенность которых заключается в наличии органов полового размножения. К ним относится большая часть всех растений на Земле — от водяных лилий и полевых цветов до дубов и фруктовых деревьев.
Автор фото, Getty Images
Подпись к фото,
В природе существует более 200 тысяч разновидностей цветковых растений
Дарвин не мог объяснить процесс их возникновения и эволюции. Цветковые растения появились на Земле сравнительно поздно по сравнению с другими видами и очень быстро приобрели широчайшее многообразие цвета, размера и форм.
«Судя по так называемой летописи ископаемых, цветковые растения (Angiospermae) появились внезапно — в меловом периоде, около 100 миллионов лет назад. У них нет ни единого сходства с растениями, существовавшими до этого. Кроме того, их появление было ознаменовано разнообразием подвидов», — говорит профессор Баггс.
У них нет ни единого сходства с растениями, существовавшими до этого. Кроме того, их появление было ознаменовано разнообразием подвидов», — говорит профессор Баггс.
Именно эта внезапность не давала покоя Чарльзу Дарвину.
Почему не было последовательной эволюции? Куда подевались промежуточные формы между хвойными породами (Gymnospermае) и цветковыми? И как это возможно, что они появились сразу в великом множестве вариантов?
Автор фото, Getty Images
Подпись к фото,
Поля тюльпанов в Германии
Дарвин не понимал, как эти растения избежали последовательных этапов развития в отличие от других обширных видов флоры и фауны, включая млекопитающих. Все это противоречило одному из главных принципов естественного отбора, который гласил, что природа не совершает резких скачков.
Долгое время Дарвин утешал себя идеей, что, возможно, цветковые растения возникли и эволюционировали на каком-нибудь еще не открытом острове или континенте.
В августе 1881 года, всего за несколько месяцев до смерти, он написал Хукеру: «Для меня нет ничего более экстраординарного в истории растительного царства, чем неожиданное и стремительное развитие высших растений. Иногда мне казалось, что на протяжении веков где-то около Южного полюса мог существовать отдаленный и затерянный континент».
Нападки Каррутерса
В библиотеке Королевских ботанических садов Кью профессор Баггс наткнулся на копию лекции, которую в 1876 году читал членам Ассоциации геологов шотландский ботаник Уильям Каррутерс.
В ней шотландец утверждает, что Дарвин не в силах понять и объяснить возникновение цветковых растений, потому что их появление имеет божественную основу.
Каррутерс обрушивается на всю дарвиновскую теорию эволюции в целом, чем провоцирует горячие споры не только в научных кругах, но и в обществе. Его высказывания и выводы напечатала газета Times, а также ряд научных изданий.
Автор фото, Getty Images
Подпись к фото,
Зал ботаники в Британском музее, 1858 год
«Каррутерс воспользовался моментом для того, чтобы начать поход против дарвиновской теории. Он утверждал, что покрытосеменные в Меловом периоде были созданы непосредственно Богом. Для Дарвина и его друзей это было полной ересью, но возникла проблема: он не мог объяснить это явление с точки зрения эволюции», — говорит Баггс.
По мнению профессора, именно эта ситуация и подтолкнула Чарльза Дарвина к использованию словосочетания «ужасная тайна». Свои выводы Баггс опубликовал в научном издании American Journal of Botany.
Сам Уильям Каррутерс впоследствии стал хранителем раздела ботаники в Британском музее и одним из крупнейших ученых в области палеоботаники.
По словам Ричарда Баггса, «ужасная тайна» Чарльза Дарвина сродни теореме Ферма, сформулированной математиком Пьером Ферма в 1637 году, — ни один из них не смог при жизни разгадать собственную загадку.
«Мы получили представление о том, что творилось в голове Дарвина в последние годы его жизни. Эта последняя загадка, попытки ее решить занимали все мысли Дарвина вплоть до самой смерти», — говорит профессор Баггс.
Удалось ли ученым с тех пор разгадать «ужасную тайну»?
Автор фото, Getty Images
Подпись к фото,
За 140 лет выдвигались различные теории происхождения цветов на Земле, но все они оставляют открытыми множество вопросов
Если одним словом, то нет.
Прошло уже 140 лет, а исчерпывающе объяснить возникновение цветковых растений до сих пор никто не может.
«Конечно, мы значительно продвинулись в нашем понимании эволюции и в знаниях палеонтологии, но эта тайна все еще не раскрыта», — говорит Ричард Баггс.
История кафедры биологии растений и методики преподавания биологии
Кафедра биологии растений и методики преподавания биологии создана в 1962 году через год после открытия факультета биологии и основ сельского хозяйства.
Кафедру основ сельского хозяйства возглавил кандидат сельскохозяйственных наук Ахмадеев Г.Х., а ботаники — кандидат сельскохозяйственных наук Белоногов Е.В., который после их объединения руководил кафедрой ботаники вплоть до 1974 года.
В числе первых преподавателей кафедры — Белоногова Л.В., Саяпова Р.Ш., Сюзева Н.В., Хамитова А.Г., Каминский А.А., Каминская А.Ф., Васильев Л.А., Калинин И.Ф., Порватова Г.Т., Гайнцева Н.С., незабвенный Г.В. Артемьев, кандидат биологических наук, блестящий оратор, с энциклопедическими знаниями биологии.
С момента открытия кафедры вплоть до выхода на заслуженный отдых преподавали: физиологию растений доцент Левченко Н.К., (первый декан факультета), микробиологию — Азимова Н.Г., основы сельского хозяйства — старший преподаватель Ахмадеев Г.Х. В 70-е годы на кафедре работали кандидаты наук Шарый М.А. (основы сельского хозяйства), Незнаев П.Д. (методика преподавания биологии), в 80-е годы — Мусина Л.С. (ботаника), доцент Мечиславский Ю. А., старший преподаватель Запольских В.Я.
А., старший преподаватель Запольских В.Я.
В течение последних 30 лет на кафедре трудились: спокойный, уравновешенный, доброжелательный к студентам доцент Шмелев А.Н. (преподавал с 1972 года), с 1975 года — доцент Кириллова Д.Ю., энергичная, умеющая поддерживать интерес к предмету, требовательная к себе и к студентам, с 1977 года после работы в Башкирском филиале Академии наук пришла на кафедру доцент Шакирова Р.Х., которую отличает солидная теоретическая подготовка, помогающая в доступной форме раскрывать один из сложнейших предметов «Физиология растений».
С 1979 года читал лекции и проводил лабораторные занятия по предмету «Ботаника» доцент Мещеряков Г.И., который являлся истинным профессионалом, любящим и знающим мир растений, активно занимающимся наукой, и признанием его значительного вклада в теоретическую науку и преподавательскую деятельность является дважды присужденная ему Премия Сороса.
На сегодняшний день на кафедре работают преподаватели, обладающие солидным багажом знаний: доценты Минина Н. Н., Рябова Т.Г., Черных И.В. (Ботаника с основами фитоценологии), доцент Трапезников доцент В.П., (Микробиология, Биологические основы сельского хозяйства), Шмелев Н.А. (Физиология растений), доцент Кириллова С.С. (Биогеография), Яппарова Э.Н. (Физиология растений), старшие преподаватели Киржакова В.Б., Талыпова Л.З., Венгилевских Ю.А. (Теория и методика обучения биологии), доцент ассистент Пурина Е.С. («Экология»).
Н., Рябова Т.Г., Черных И.В. (Ботаника с основами фитоценологии), доцент Трапезников доцент В.П., (Микробиология, Биологические основы сельского хозяйства), Шмелев Н.А. (Физиология растений), доцент Кириллова С.С. (Биогеография), Яппарова Э.Н. (Физиология растений), старшие преподаватели Киржакова В.Б., Талыпова Л.З., Венгилевских Ю.А. (Теория и методика обучения биологии), доцент ассистент Пурина Е.С. («Экология»).
Хочется вспомнить о важнейших помощниках наших преподавателей — лаборантах. Много лет работали на кафедре Кирюхина В.Л., Репина М.А., Смородинова Ф.Н., Гелимьянова З.М., Сабанцева А.Г., несколько лет работали лаборантами Анисимова Е.Н., Выдрина Н.М., Мальцева Г.Н. Долгие годы была старшим лаборантом Самойлова Т.С. Учебными мастерами работали Хахалкина В.М., Габбасов Р.Р.
В настоящее время коллектив лаборантов возглавляет показавшая умение работать Иманова Э.А., в числе лаборантов Агафонова Т.А., Зинова Л.А. Агростанцию возглавляет Чуклина О. В. учебный мастер — Валиахмедов Б.В.
В. учебный мастер — Валиахмедов Б.В.
История комнатных растений
История комнатных растений
Как комнатные растения пришли в наш дом ?
Комнатные растения так прочно вошли в нашу жизнь, что трудно уже представить себе дом без привычных горшков на подоконнике, офис без неизменного фикуса в кадке или холл общественного учреждения без композиции из нескольких таких «фикусов».
Многие ласково именуют их «домашними» (видимо, по аналогии с домашними животными), не задумываясь о том, что «приручение» растений состоялось сравнительно недавно.
Впрочем, в какой-то мере историю комнатных растений можно начинать еще с Древнего Египта, где цветы высаживались в просторные кадки и украшали жилища и общественные здания. На древнеегипетских изображениях, датируемых более чем тысячелетием до н.э , хорошо различимы маленькие деревья и кустики в каменных вазах и лотках. Но эти растения еще не были в полной мере «комнатными», хотя, пожалуй, могли уже считаться «домашними».
Но эти растения еще не были в полной мере «комнатными», хотя, пожалуй, могли уже считаться «домашними».
Сейчас нам, в любом цветочном магазине покупающим тропических экзотов, родившихся и выросших в горшках, трудно вообразить, какой огромный путь лежит между первым привезенным из далеких земель ростком и нынешним огромным ассортиментом.
Эпоха великих географических открытий, случившаяся в истории человечества в XV веке, стала весьма значительным этапом и во взаимоотношениях человека с растениями. Путешественники из Европы, покоряя новые земли, были потрясены красотой и многообразием тропических растений. Семена, черенки, листочки и растения целиком вывозились на родину, где их, из-за невозможности адаптировать к более суровому климату, выращивали в помещениях.
Именно таким образом в Старом Свете и появилось большинство комнатных растений. Эти растения тут стали предметом гордости, символом роскоши и престижа. Их холили, лелеяли и берегли не ради «общения» с ними, а исключительно в качестве «диковинок», призванных поражать гостей и служить атрибутом исключительности хозяев. Впрочем, по мере распространения комнатных растений и приобретения опыта ухода за ними, вырванные из привычной среды обитания экзоты перестали быть чем-то необычным и прочно обрели новую форму жизни в доме человека.
Впрочем, по мере распространения комнатных растений и приобретения опыта ухода за ними, вырванные из привычной среды обитания экзоты перестали быть чем-то необычным и прочно обрели новую форму жизни в доме человека.
Говоря об истории комнатных растений, невозможно не упомянуть зимние сады. Устройство первого «настоящего» зимнего сада приписывают Альберту Магнусу, философу, теологу и ученому, жившему в XIII веке. Мне не удалось найти достоверных сведений о подобном проекте Альберта Великого, но, несомненно, этот человек достоин упоминания в связи с растениями, поскольку он был выдающимся для своего времени ботаником, автором научного труда De vegetalibus et plantis (О растениях). В честь Альберта Великого даже назван род африканских деревьев с красивыми трубчатыми цветками из семейства Мареновые — Альберта (Alberta).
Прообразами зимних садов в Европе XVI столетия стали так называемые «дома померанцев». Это были теплицы, в которых выращивались экзотические растения и плоды, в первую очередь апельсины (по-французски «orange»), откуда и пошло название «оранжерея». Первые оранжереи были построены в Ботаническом саду Лейдена в 1599 году, в 1646 году они появились в Ренте и Амстердаме, в 1714 — в Париже. Одновременно с парижской появилась и первая русская оранжерея, построенная по инициативе Петра I в Петербурге в Летнем саду. В XVIII веке многие знатные семьи обеспечивали себя экзотическими фруктами и овощами за счет оранжерей при своих городских и загородных усадьбах. Размах оранжерейного дела тех времен способен и сегодня вызвать завистливое изумление.
Первые оранжереи были построены в Ботаническом саду Лейдена в 1599 году, в 1646 году они появились в Ренте и Амстердаме, в 1714 — в Париже. Одновременно с парижской появилась и первая русская оранжерея, построенная по инициативе Петра I в Петербурге в Летнем саду. В XVIII веке многие знатные семьи обеспечивали себя экзотическими фруктами и овощами за счет оранжерей при своих городских и загородных усадьбах. Размах оранжерейного дела тех времен способен и сегодня вызвать завистливое изумление.
В XIX веке городские усадьбы уступили место особнякам, и растения стали по-настоящему «комнатными», переместившись внутрь жилых помещений. В России в этот период уже были широко известны такие комнатные растения, как пальмы, фикус, абелия, гибикус китайский, монстера, пеларгония, различные цитрусовые.
Невероятно важным для комнатного цветоводства оказалось открытие англичанина Н.Уорда, который заметил, что в небольшой стеклянной камере, по причине высокой влажности воздуха, даже нежные папоротники чувствуют себя вполне удовлетворительно. В 1834 году Уорд сконструировал первый «чемодан» для перевозки растений, который устанавливали на палубе корабля. Растениям требовалось меньше полива и они были защищены от соленых брызг. Теперь, благодаря приспособлениям, вошедшим в историю под названием «ящики Уорда», в Европу без потерь можно было привозить нежные тропические растения: папоротники, орхидеи, бромелиевые.
В 1834 году Уорд сконструировал первый «чемодан» для перевозки растений, который устанавливали на палубе корабля. Растениям требовалось меньше полива и они были защищены от соленых брызг. Теперь, благодаря приспособлениям, вошедшим в историю под названием «ящики Уорда», в Европу без потерь можно было привозить нежные тропические растения: папоротники, орхидеи, бромелиевые.
Это изобретение позволило не только увеличить приток экзотических растений в страны Старого Света, но и расширить их ассортимент за счет более прихотливых культур. В наши дни производство комнатных растений — целая отрасль цветоводства. С одной стороны, мы теперь имеем возможность без особых трудов приобрести практически любой комнатный цветок, с другой — полностью утрачен ореол романтики и новизны, окружавший эти растения в минувшие столетия. Став расхожим товаром, выходцы из тропиков, некогда поражавшие воображение людей, теперь скромно занимают место «в уголке» нашей жизни. Но и здесь они незаметно выполняют свою великую роль. ..
..
Из книги Юлии Фоминой «Красивое комнатное цветоводство»
Еще об истории комнатных растений
Гарден Ритейл Сервис — лидер рынка производства грунтов и удобрений для сада и огорода
Производственно-складской комплекс «Снопово»
Производственно-складской комплекс «Снопово» европейского уровня класса «А» – это современное передовое оборудование и самые уникальные технологии производства. ПСК «Снопово» работает с 1993 года.
Для производства лучших грунтов на рынке на территории комплекса установлена высокотехнологичная линия по глубокой переработке торфа и автоматизированная фасовочная линия.
Данное оборудование дает возможность:
- Отгружать грунты и торфяную продукцию в течении 48 часов с момента размещения заказа.
- Производить и разрабатывать продукты с дополнительными компонентами, такими как Гидрогель, Биогумус и др.

- Контролировать точное весовое и объемное дозирование, а также другие параметры выпускаемых продуктов на всех этапах производства.
- Выпускать готовую продукцию во всех востребованных фасовках от 1 литра до 100 литров.

Производственно-складской комплекс «Снопово» каждый день может производить:
- Более 1200м3 грунтов и торфяной продукции;
- Более 20 тонн минеральных удобрений;
- Более 9000 литров жидких концентрированных удобрений;
- Более 110 000 штук средств защиты растений.
Ежедневные отгрузки в сезон составляют от 500 до 900 тонн в сутки:
- 40 и более грузовых а/м МО;
- 30 и более крупнотоннажных грузовых а/м России и СНГ;
- До 3-х вагонов РФ и СНГ.
Собственный отдел качества ежедневно осуществляет строгий контроль отгружаемой продукции.
Зона уличного хранения рассчитана на 20 000 паллет.
Годовой грузооборот склада составляет более 110 000 тонн.
Собственный торфяник в экологически чистом регионе Владимирской Области
В 2016 году компании была выдана лицензия на разведку и добычу торфа в Гусь-Хрустальном районе. За два года была проведена оценка запасов торфа на Иванищевском и Суловско-Панфиловском участках, подготовлены проектные решения по разработке и рекультивации месторождения торфа, выбрана площадка для строительства завода по глубокой переработке торфа c высоким уровнем автоматизации.
На сегодня плановый объём добычи торфа составляет свыше 400 000 м3 в год.
На торфянике и производстве задействовано около 200 человек. На заводе выпускается продукция по уходу за уличными и комнатными растениями.
Собственные питомники растений в Московской области и Крыму
«Тимирязевский питомник. Филиал Северный» и «Лаванда. Филиал Южный» — это два питомника по взращиванию растений, в которых представлен широкий выбор плодовых и хвойных растений, декоративных кустарников и цветов, которые придутся по вкусу самому взыскательному покупателю.
Агрономы и специалисты проходят регулярные обучения и стажировки в ведущих Аграрных университетах. Увлеченные своим делом, они с любовью и трепетом относятся к каждому выращиваемому растению. Благодаря передовым технологиям, опыту и знаниям профессиональных сотрудников, растения легко приживутся на садовых участках будущих владельцев, будут радовать глаз высокой декоративностью, доставлять наслаждение высокими урожаями.
Преимущества питомников:
- Современные теплицы с высокотехнологичным оборудованием;
- Применение опыта ведущих европейских производителей;
- Наличие собственных маточников, что позволяет выращивать необходимое количество растений по лидерской цене;
- Полный производственный цикл, что исключает зависимость от курса валют и государственных санкций;
- Приоритет в пользу качества растений перед массовым объемом;
- Гибкий и клиентоориентированный подход, возможность выращивать растения под потребности заказчика (сорт, размер).

«Тимирязевский питомник. Филиал Северный» был основан в 2015 году на территории 2 Га в Солнечногорском районе Московской области. В настоящее время оборудован 12 современными теплицами по 400 кв. м. каждая. Ежегодно техническое оснащение питомника дополняется и обновляется согласно последним европейским тенденциям и технологиям.
«Тимирязевский питомник. Филиал Северный» осуществляет:
- Полный цикл выращивания садовой земляники и горшечных растений и рассады;
- Доращивание растений, акклиматизацию и закаливание теплолюбивых культур;
- Производство зимостойкого посадочного материла.
Качество посадочного материала является исключительным для выращивания в Москве и Московской области, что служит гарантом максимальной адаптации и приспособленности растений к условиям дальнейшего произрастания.
Марка «Тимирязевский питомник. Филиал Северный» говорит о том, что:
- Растение было выращено в соответствии с высокими стандартами марки;
- Тщательно отобрано для климатической зоны Вашего сада;
- Тщательно протестировано перед продажей;
- Растения поступают в продажу только здоровыми.

Крепкие, здоровые растения с опережающим ровесников ростом! Наши контейнеры для растений оптимально подобраны с учетом роста и развития конкретного вида растения, наш почвогрунт — профессиональная смесь. Мы не экономим на материале. Это позволяет растениям развиваться с большей корневой системой для здорового роста. Это означает, что Вы получите устоявшееся растение, которое не будет испытывать стресс при пересадке.
Все растения выращиваются только с учетом особенностей климата нашей территории и не нуждаются в акклиматизации. Это означает, что ваши растения благополучно перенесут морозы нашей климатической зоны.
Акклиматизированные растения с мощной корневой системой, выглядят более «взросло, не испытывают стресс при пересадке, готовы «взяться за дело» с уверенным ростом. Вы несомненно будете удовлетворены результатом! Адаптированные к нашей территории и профессионально выращенные растения с мощной корневой системой обеспечат вам ранний и обильный урожай.
Все растения перед продажей тестируются повторно.
Агрономы ТИМИРЯЗЕВСКОГО ПИТОМНИКА филиал северный хорошо знают свое дело и трудятся ежедневно ради достижения самых высоких результатов. Каждое растение, выращивается под тщательным контролем специалистов питомника, внимательным образом тестируется и поступает в продажу только здоровым. Яблоня, груша, виноград, малина, жимолость, слива, смородина и многие.
«Лаванда. Филиал Южный» – это современные теплицы с высокотехнологичным оборудованием, собственные маточники и большой выбор растений. История создания Питомника «Лаванда» началась в 2018 году, когда летом стартовали работы по постройке современных теплиц на территории 8 Га в Бахчисарайском районе солнечного Крыма.
Питомник Лаванда – это полный производственный цикл, основанный на современных технологиях. Растения районированы и подходят для большинства климатических зон РФ.
Собственная лаборатория по разработке продукции
Собственная лаборатория компании «Гарден Ритейл Сервис» позволяет разрабатывать новые продукты и поддерживать качество выпускаемых на самом высоком уровне.
Многие изделия производятся по уникальным сбалансированным рецептурам, не имеющим аналогов — ученые находят идеально подобранные формулы макро- и микроэлементов, позволяющие добиться высокой эффективности. Вместе с тем, специалисты компании учитывают большой опыт, накопленный за 25 лет ответственной работы.
Прежде чем выпускать новую продукцию на рынок, проводятся многочисленные тесты и исследования — все разработанные продукты обеспечивают повышение урожайности и способствуют гармоничному развитию растений.
Вся продукция имеет необходимые сертификаты и включена в список разрешенных к применению препаратов.
The Greenville News
Durant Ashmore
| Специально для The Greenville News
Чем больше вы знаете о растениях, тем больше удовольствия они приносят. Что я обнаружил недавно, так это то, что растения, которые мы используем в наши дни, имеют невероятно широкий диапазон исторического происхождения.
Модные садоводы любят использовать новинки в модных садовых растениях, и я это понимаю. Приятно пробовать новое. Однако по мере того, как мой садоводство становится «зрелым», меня привлекают некоторые из более древних растений — и в сегодняшних примерах я имею в виду действительно, очень древние.
Приятно пробовать новое. Однако по мере того, как мой садоводство становится «зрелым», меня привлекают некоторые из более древних растений — и в сегодняшних примерах я имею в виду действительно, очень древние.
Мне нравится изучать историю вещей. В сегодняшней колонке мы поговорим об истории растений.
Есть некоторые свидетельства — хотя и отсутствуют ископаемые остатки — того, что первые растения произошли от зеленых водорослей в пресноводных прудах 1 000 миллионов лет назад. Эти растения вымерли сотни миллионов лет назад.
Самыми старыми известными растениями, живущими сегодня, являются печеночники. Почти на любой суше в мире произрастает 9000 видов печеночников.Самые ранние печеночники существуют в летописи окаменелостей 470 миллионов лет назад.
Следующими в летописи окаменелостей появляются мхи. В мире насчитывается 14 500 видов мха. Есть даже ископаемые остатки мха, найденные в Антарктиде примерно 275 миллионов лет назад.
Печеночники и мхи классифицируются как бриофиты.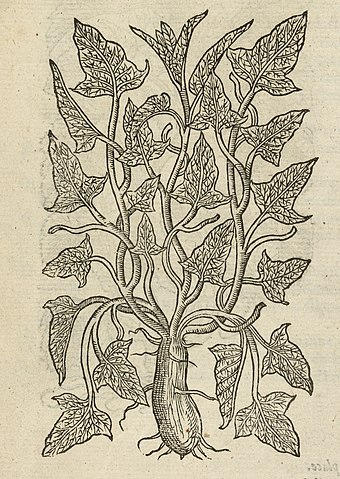 Эти растения не имеют сосудистой системы и не могут проводить воду или другие питательные вещества по всему растению.
Эти растения не имеют сосудистой системы и не могут проводить воду или другие питательные вещества по всему растению.
Как вы понимаете, сосудистая система была бы важным эволюционным преимуществом растения.Вот где в игру вступают папоротники. 12 000 видов папоротников впервые вошли в летопись окаменелостей 360 миллионов лет назад. В течение миллионов лет папоротники были доминирующим растением в мире, причем папоротниковые леса достигали 80 футов в высоту.
Папоротники, мхи и печеночники размножаются спорами, что является одной из форм бесполого размножения. Следующий большой шаг в эволюции растений происходит с половым размножением, когда пыльца играет важную роль.
Первыми растениями, размножающимися половым путем, являются голосеменные, происходящее от греческого слова, означающее «голое семя».Вы всю жизнь знакомы с голосеменными. Сосны, как и все другие хвойные породы, являются голосеменными. Голосеменные впервые вошли в летопись окаменелостей около 315 миллионов лет назад.
Однако хвойные растения — не единственные голосеменные, с которыми мы знакомы в повседневной жизни. Древние деревья гинкго и подокарпусы также являются голосеменными.
Гинкго — самые старые живые деревья, зарегистрированные в летописи окаменелостей. Они существуют уже 65 миллионов лет. Они почти вымерли, и лишь несколько ничтожных экземпляров жили в горах Китая, прежде чем современное садоводство спасло их от забвения.Теперь, благодаря яркому изображению великолепного золотого осеннего цвета, деревья гинкго становятся любимой частью любого пейзажа.
Подокарпус также является ценным представителем современного ландшафта. Хотя Пьемонт — самый северный ареал этого растения, его использование в ландшафте добавляет характер любой обстановке. Нет, не цветет (это голосеменное растение, запомните). Однако текстура подокарпуса непревзойденная. Признание фактуры пейзажа — еще один способ эволюции вкусов зрелого пейзажа.
С появлением цветковых растений 140 миллионов лет назад развились покрытосеменные. Покрытосеменные переводятся с греческого как «семя, рожденное в сосуде». У покрытосеменных есть инкапсулированные семена, такие как фрукты и орехи.
Покрытосеменные переводятся с греческого как «семя, рожденное в сосуде». У покрытосеменных есть инкапсулированные семена, такие как фрукты и орехи.
Самыми древними цветущими растениями, сохранившимися до наших дней, являются кувшинки. Древние цветковые растения характеризуются большими, открытыми, эффектными одиночными цветками на каждом растении. Магнолия — еще одно древнее растение, которое демонстрирует эту характеристику, и на самом деле покрытосеменные растения также называют «магнолияфитами».
Вы можете сказать, как далеко в древней истории возникло растение, взглянув на его цветочную структуру.Например, посмотрите на анис Флориды. У этого растения, произрастающего во Флориде, есть большой эффектный красный цветок с открытой структурой в форме звезды. Он растет в той же среде обитания, что и желтый анис, у которого во время цветения появляется множество маленьких желтых цветков. Однако флоридский анис возник намного раньше, чем желтый анис, и вы знаете это из-за цветочной структуры.
Квансанская вишня с крупными одиночными цветками возникла раньше, чем вишня Ёсино, у которой многоцветие. Фактически, любое растение, которое имеет обилие мелких цветков, является поздней версией любого растения с большими одиночными цветками. Camellia japonica (сейчас цветущая) старше Camellia sasanqua .
Все эти новомодные растения действительно служат своей цели в сегодняшнем ландшафте. Однако давайте не будем забывать о характере и стойкости более древних образцов, которые добавляют реальную ценность нашим современным ландшафтам.
завод | Определение, эволюция, экология и систематика
растение , (царство Plantae), любая многоклеточная эукариотическая форма жизни, характеризующаяся (1) фотосинтетическим питанием (характеристика, присущая всем растениям, за исключением некоторых паразитических растений и подземных орхидей), у которых химическая энергия производится из воды, минералов и углекислого газа с помощью пигментов и лучистой энергии Солнца, (2) практически неограниченный рост в локализованных областях, (3) клетки, которые содержат целлюлозу в своих стенках и поэтому в некоторой степени степень жесткости, (4) отсутствие органов передвижения, приводящее к более или менее стационарному существованию, (5) отсутствие нервной системы, и (6) истории жизни, которые показывают смену гаплоидных и диплоидных поколений с преобладанием один над другим таксономически значим.
Британская викторина
(Не) Все в семье
У растений тоже есть семьи! Постарайтесь выяснить, какие растения не принадлежат этим знаменитым семействам.
Размеры растений варьируются от миниатюрных ряски длиной всего несколько миллиметров до гигантских калифорнийских секвой, достигающих в высоту 90 метров (300 футов) и более.По оценкам, науке известно 390 900 различных видов растений, и постоянно описываются новые виды, особенно из ранее неизведанных тропических областей мира. Растения произошли от водных предков и впоследствии мигрировали по всей поверхности Земли, населяя тропические, арктические, пустынные и альпийские регионы. Некоторые растения вернулись в водную среду обитания либо в пресной, либо в соленой воде.
Растения играют жизненно важную роль в поддержании жизни на Земле. Вся энергия, используемая живыми организмами, зависит от сложного процесса фотосинтеза, который в основном осуществляется зелеными растениями. Лучистая энергия Солнца преобразуется в органическую химическую энергию в форме сахаров посредством фундаментальной серии химических реакций, составляющих фотосинтез. В природе все пищевые цепи начинаются с фотосинтетических автотрофов (первичных продуцентов), включая зеленые растения и водоросли. Первичные продуценты, представленные деревьями, кустарниками и травами, являются богатым источником энергии в виде углеводов (сахаров), хранящихся в листьях.Эти углеводы, производимые в процессе фотосинтеза, расщепляются в процессе, называемом дыханием; меньшие единицы молекулы сахара и его продуктов питают многочисленные метаболические процессы. Различные части растения (например, листья) являются источниками энергии, поддерживающими жизнь животных в различных средах обитания. Побочный продукт фотосинтеза, кислород, необходим животным.
Вся энергия, используемая живыми организмами, зависит от сложного процесса фотосинтеза, который в основном осуществляется зелеными растениями. Лучистая энергия Солнца преобразуется в органическую химическую энергию в форме сахаров посредством фундаментальной серии химических реакций, составляющих фотосинтез. В природе все пищевые цепи начинаются с фотосинтетических автотрофов (первичных продуцентов), включая зеленые растения и водоросли. Первичные продуценты, представленные деревьями, кустарниками и травами, являются богатым источником энергии в виде углеводов (сахаров), хранящихся в листьях.Эти углеводы, производимые в процессе фотосинтеза, расщепляются в процессе, называемом дыханием; меньшие единицы молекулы сахара и его продуктов питают многочисленные метаболические процессы. Различные части растения (например, листья) являются источниками энергии, поддерживающими жизнь животных в различных средах обитания. Побочный продукт фотосинтеза, кислород, необходим животным.
фотосинтез
Схема фотосинтеза, показывающая, как вода, свет и углекислый газ поглощаются растением с образованием кислорода, сахара и большего количества углекислого газа.
Encyclopædia Britannica, Inc.
Растения также напрямую влияют на повседневное существование человека. Растения снабжают пищей и ароматизаторами; сырье для промышленности, такое как древесина, смолы, масла и резина; волокна для изготовления тканей и снасти; лекарства; инсектициды; и топливо. Более половины населения Земли использует рис, кукурузу (кукурузу) и пшеницу в качестве основного источника пищи. Помимо своей коммерческой и эстетической ценности, растения сохраняют другие природные ресурсы, защищая почвы от эрозии, контролируя уровень и качество воды и создавая благоприятную атмосферу.
Получите подписку Britannica Premium и получите доступ к эксклюзивному контенту.
Подпишитесь сейчас
В следующей статье обобщаются морфологические, физиологические и экологические особенности растений. Основное внимание уделяется структуре и функциям, физиологии, истории жизни и экологии, а также тому, как различные группы растений эволюционировали, рассредоточились и стали адаптироваться к жизни на суше. Также обсуждаются особенности, которые определяют каждую основную группу растений, и роль, которую они играют в более широкой экосистеме.
Также обсуждаются особенности, которые определяют каждую основную группу растений, и роль, которую они играют в более широкой экосистеме.
завод | Определение, эволюция, экология и систематика
растение , (царство Plantae), любая многоклеточная эукариотическая форма жизни, характеризующаяся (1) фотосинтетическим питанием (характеристика, присущая всем растениям, за исключением некоторых паразитических растений и подземных орхидей), у которых химическая энергия производится из воды, минералов и углекислого газа с помощью пигментов и лучистой энергии Солнца, (2) практически неограниченный рост в локализованных областях, (3) клетки, которые содержат целлюлозу в своих стенках и поэтому в некоторой степени степень жесткости, (4) отсутствие органов передвижения, приводящее к более или менее стационарному существованию, (5) отсутствие нервной системы, и (6) истории жизни, которые показывают смену гаплоидных и диплоидных поколений с преобладанием один над другим таксономически значим.
Британская викторина
Растения: факт или вымысел?
Могут ли плотоядные растения есть людей? Насколько велико самое большое водное растение? Зеленый и черный чай изготавливают из разных растений? Искорените ответы, покопавшись в этой викторине.
Размеры растений варьируются от миниатюрных ряски длиной всего несколько миллиметров до гигантских калифорнийских секвой, достигающих в высоту 90 метров (300 футов) и более.По оценкам, науке известно 390 900 различных видов растений, и постоянно описываются новые виды, особенно из ранее неизведанных тропических областей мира. Растения произошли от водных предков и впоследствии мигрировали по всей поверхности Земли, населяя тропические, арктические, пустынные и альпийские регионы. Некоторые растения вернулись в водную среду обитания либо в пресной, либо в соленой воде.
Растения играют жизненно важную роль в поддержании жизни на Земле.Вся энергия, используемая живыми организмами, зависит от сложного процесса фотосинтеза, который в основном осуществляется зелеными растениями. Лучистая энергия Солнца преобразуется в органическую химическую энергию в форме сахаров посредством фундаментальной серии химических реакций, составляющих фотосинтез. В природе все пищевые цепи начинаются с фотосинтетических автотрофов (первичных продуцентов), включая зеленые растения и водоросли. Первичные продуценты, представленные деревьями, кустарниками и травами, являются богатым источником энергии в виде углеводов (сахаров), хранящихся в листьях.Эти углеводы, производимые в процессе фотосинтеза, расщепляются в процессе, называемом дыханием; меньшие единицы молекулы сахара и его продуктов питают многочисленные метаболические процессы. Различные части растения (например, листья) являются источниками энергии, поддерживающими жизнь животных в различных средах обитания. Побочный продукт фотосинтеза, кислород, необходим животным.
фотосинтез
Схема фотосинтеза, показывающая, как вода, свет и углекислый газ поглощаются растением с образованием кислорода, сахара и большего количества углекислого газа.
Encyclopædia Britannica, Inc.
Растения также напрямую влияют на повседневное существование человека. Растения снабжают пищей и ароматизаторами; сырье для промышленности, такое как древесина, смолы, масла и резина; волокна для изготовления тканей и снасти; лекарства; инсектициды; и топливо. Более половины населения Земли использует рис, кукурузу (кукурузу) и пшеницу в качестве основного источника пищи. Помимо своей коммерческой и эстетической ценности, растения сохраняют другие природные ресурсы, защищая почвы от эрозии, контролируя уровень и качество воды и создавая благоприятную атмосферу.
Получите подписку Britannica Premium и получите доступ к эксклюзивному контенту.
Подпишитесь сейчас
В следующей статье обобщаются морфологические, физиологические и экологические особенности растений. Основное внимание уделяется структуре и функциям, физиологии, истории жизни и экологии, а также тому, как различные группы растений эволюционировали, рассредоточились и стали адаптироваться к жизни на суше. Также обсуждаются особенности, которые определяют каждую основную группу растений, и роль, которую они играют в более широкой экосистеме.
Основное внимание уделяется структуре и функциям, физиологии, истории жизни и экологии, а также тому, как различные группы растений эволюционировали, рассредоточились и стали адаптироваться к жизни на суше. Также обсуждаются особенности, которые определяют каждую основную группу растений, и роль, которую они играют в более широкой экосистеме.
Функциональные черты объясняют различия в стратегиях жизненного цикла растений
Значимость
Растения выработали разнообразные стратегии жизненного цикла, чтобы добиться успеха в разнообразных средах Земли. Некоторые виды быстро растут, дают обильные семена и погибают в течение нескольких недель. Другие виды растут медленно и редко дают семена, но живут тысячи лет. Мы показываем, что простые морфологические измерения могут предсказать, где вид попадает в глобальный диапазон стратегий жизненного цикла: у видов с крупными семенами, долгоживущими листьями или густой древесиной на темпы роста популяции влияет в первую очередь выживание, тогда как индивидуальный рост и плодовитость имеют более сильное влияние на динамику видов с мелкими семенами, короткоживущими листьями или мягкой древесиной.Это открытие увеличивает способность ученых представлять сложные популяционные процессы с помощью нескольких легко измеряемых черт характера.
Abstract
Экологи ищут общие объяснения резкого изменения численности видов в пространстве и времени. Все более популярным решением является прогнозирование распределения видов, динамики и реакции на изменение окружающей среды на основе легко измеряемых анатомических и морфологических признаков. Подходы, основанные на признаках, предполагают, что простые функциональные признаки влияют на приспособленность и эволюцию жизненного цикла, но строгие проверки этого предположения отсутствуют, поскольку они требуют количественной информации о полных жизненных циклах многих видов, представляющих различные жизненные истории.Здесь мы связываем глобальную базу данных по признакам с эмпирическими матричными моделями популяций для 222 видов и сообщаем о тесной взаимосвязи между функциональными признаками и историей жизни растений. Виды с крупными семенами, долгоживущими листьями или густой древесиной имеют медленную историю жизни, при этом средняя приспособленность (т. Е. Темпы роста популяции) в большей степени зависит от выживания, чем от роста или плодовитости, по сравнению с видами с быстрой историей жизни с небольшими семенами, короткими живые листья или мягкая древесина. В отличие от мер демографического вклада в приспособленность на основе полных жизненных циклов, анализ, сфокусированный на первичных демографических показателях, может недооценивать силу связи между характеристиками и средней приспособленностью.Наши результаты помогают установить физиологическую основу эволюции жизненного цикла растений и показать потенциал подходов, основанных на признаках, в динамике популяций.
Недавние данные о глобальных закономерностях функциональной изменчивости растений, такие как экономический спектр листа (1, 2), экономический спектр древесины (3) и компромисс между размером семян и количеством семян (4, 5), убедили многих Экологи считают, что функциональные особенности предлагают наилучший доступный подход для достижения общего прогнозного понимания сообществ и экосистем (6, 7).Подходы, основанные на признаках, в настоящее время используются для прогнозирования результатов объединения сообществ (8⇓ – 10), глобальной динамики растительности (11) и скорости экосистемных процессов (6, 12⇓ – 14). Основное предположение экологии, основанной на признаках, заключается в том, что морфологические признаки определяют физиологические характеристики, которые влияют на показатели жизнедеятельности и определяют индивидуальную приспособленность и эволюцию жизненного цикла (15, 16). Однако из-за проблемы количественной оценки вклада черт в приспособленность предполагаемые связи между функциональными чертами и историей жизни не были полностью проверены.
Исследования в тропических и средиземноморских лесах выявили межвидовые связи между функциональными признаками, выживаемостью и темпами роста особей (3, 17⇓⇓⇓⇓⇓⇓ – 24). Хотя эти отношения свидетельствуют о том, что функциональные черты влияют на показатели жизнедеятельности, они дают лишь ограниченное представление о связях между этими чертами и индивидуальной приспособленностью и историей жизни. Показатели жизнеспособности (например, выживаемость и плодовитость) представляют собой компоненты приспособленности, но их влияние на среднюю приспособленность, определяемую как скорость роста популяции (λ), лучше всего понимать в контексте полного жизненного цикла вида (25, 26).Существенная отрицательная корреляция между плотностью древесины и индивидуальным ростом (18) может не привести к значительному влиянию на среднюю приспособленность, если индивидуальный рост мало влияет на λ. И наоборот, слабая связь между функциональным признаком и другим показателем жизнеспособности может иметь значительное влияние на среднюю приспособленность, если этот показатель жизнеспособности имеет сильное влияние на λ. Анализ возмущений, таких как чувствительность и эластичность, часто применяемый к матричным проекционным моделям (27), решает эту проблему путем количественной оценки вклада показателей жизнедеятельности в λ (28), что позволяет охарактеризовать общую историю жизни вида с точки зрения относительная важность выживания, индивидуального роста и плодовитости для обозначения приспособленности.Виды с медленным жизненным циклом имеют темпы роста популяции с высокой эластичностью к выживанию, тогда как виды с быстрым жизненным циклом имеют относительно более высокую эластичность к индивидуальному росту или плодовитости (29, 30).
Вооружившись эластичностью показателей естественного движения населения, мы можем проверить количественные гипотезы о том, могут ли функциональные компромиссы увеличиваться, чтобы генерировать компромиссы для истории жизни. Например, растения могут направить свои репродуктивные усилия на обеспечение нескольких крупных семян, которые выдерживают слабое освещение и доступность ресурсов и имеют высокую вероятность выживания, или они могут распределить свои репродуктивные усилия между множеством мелких семян, максимизируя приспособленность при высокой доступности ресурсов (31 , 32).Если этот функциональный компромисс на стадии проростков трансформируется в компромисс между жизненным циклом, масса семян должна быть положительно связана с эластичностью темпа роста популяции по отношению к выживаемости и отрицательно связана с эластичностью по индивидуальному росту и плодовитости. Спектр листовой экономики представляет собой еще один компромисс при распределении ресурсов. Виды могут образовывать долгоживущие, хорошо защищенные листья, которые часто используются в условиях ограниченных ресурсов, или строить листья, которые быстро усваивают углерод в условиях высокой доступности ресурсов, но склонны к быстрой потере тканей (1, 33).Виды с медленными экономическими характеристиками листьев, такими как большая продолжительность жизни листа, низкая удельная площадь листа (SLA) и низкий уровень азота листа, также могут вести медленный образ жизни, характеризующийся высокой эластичностью к выживанию и низкой эластичностью к индивидуальному росту и пополнению. Также существует спектр экономики древесины: виды с густой древесиной, как правило, имеют более высокую выживаемость, но более низкие относительные темпы роста, чем виды с мягкой древесиной (3, 34). Эластичность для выживания должна увеличиваться с увеличением плотности древесины, тогда как эластичность для индивидуального роста и плодовитость должны уменьшаться.
Основным препятствием при проверке этих гипотез является доступность подробных демографических данных, необходимых для описания полного жизненного цикла вида и оценки эластичности жизненного цикла. Мы преодолели это ограничение, скрестив Глобальную базу данных признаков растений TRY (35) с Матричной базой данных растений COMPADRE (www.compadre-db.org/), коллекцией опубликованных матричных моделей популяций. Этот подход позволил получить набор данных из 222 видов растений, охватывающих глобальный диапазон биомов и многолетних форм роста (таблица S1), для которых у нас есть по крайней мере одно измерение функциональных признаков, а также матричная популяционная модель, которую мы использовали для расчета эластичности темпы роста населения к каждому из трех показателей жизнедеятельности: выживаемости, росту и плодовитости (30).
Наша основная цель состояла в том, чтобы оценить способность функциональных признаков объяснять различия между видами в истории жизни, которую мы количественно оценили с помощью эластичности жизненного цикла. Нашей вторичной целью было оценить, можно ли сделать выводы об истории жизни непосредственно из необработанных показателей естественного движения населения, что сэкономило бы исследователям значительное время и усилия, необходимые для параметризации моделей населения и расчета эластичности. Мы использовали два статистических подхода для количественной оценки взаимосвязи между эластичностью жизненного цикла и массой семян, плотностью древесины и экономическими характеристиками листьев (продолжительность жизни листа, SLA и N листа).Регрессия Дирихле — это многомерный подход, который учитывает тот факт, что эластичности выживания, роста и плодовитости для каждого вида в сумме равны единице, но не учитывают филогенетические отношения. Филогенетическая регрессия обобщенных наименьших квадратов (PGLS) игнорирует независимость эластичностей, но учитывает филогенетические отношения. Мы повторили оба типа регрессии с формой роста растений, а затем включили биом в качестве ковариатов, чтобы подтвердить, что эффекты признаков не просто отражают различия между деревьями и видами трав или растениями, адаптированными к разным условиям.
Результаты
Мы нашли убедительную поддержку гипотезы о том, что виды с медленными анатомическими и морфологическими признаками также имеют медленную историю жизни, которая измеряется эластичностью темпа роста популяции по каждому из трех показателей жизнедеятельности (рис. 1, рис. S1 и таблицы S2 – S5). Как и предполагалось, виды с большей массой семян имели более высокую эластичность к выживанию как в регрессии Дирихле, так и в регрессии PGLS, даже с формой роста и биомом, включенными в качестве ковариатов. В регрессии Дирихле эластичность к индивидуальному росту и плодовитость не зависела от массы семян.Эластичность выживания увеличивается с плотностью древесины; эта взаимосвязь была чувствительна к включению формы роста в регрессию Дирихле, но не к регрессии PGLS. Регрессии Дирихле не выявили значимых взаимосвязей между плотностью древесины и эластичностью индивидуального роста или плодовитости. Виды с более долгоживущими листьями также характеризовались более высокой эластичностью к выживанию. Регрессия Дирихле показала, что этот эффект отражает различия в форме роста. Однако, когда мы удалили один очевидный выброс ( Pinus nigra , вид с длинной продолжительностью жизни листа, но с очень низкой эластичностью при выживании, о которой сообщалось в быстро расширяющейся инвазивной популяции) (36), продолжительность жизни листа была положительно связана с эластичностью к выживанию, даже при наличии ростовой формы ( Z = 2.8, P = 0,006) (рис. S2). Добавление биома в качестве ковариаты не оказало качественного влияния на эти отношения. Хотя масса семян, плотность древесины и продолжительность жизни листьев в значительной степени коррелировали с эластичностью выживания, эти признаки объясняли лишь небольшую часть вариаций. В простых линейных регрессиях логит-преобразованных эластичностей выживаемости R 2 составляло 0,24, 0,08 и 0,29 для массы семян, плотности древесины и продолжительности жизни листьев, соответственно.
Рис. 1.
Функциональные характеристики связаны с эластичностью жизненного цикла.Взаимосвязи оценивались с помощью регрессии Дирихле. Пунктирными линиями показаны незначительные эффекты функциональных признаков, сплошными линиями показаны значительные эффекты функциональных признаков, а пунктирными линиями показаны эффекты функциональных признаков, которые не были значительными после того, как в модель была добавлена форма роста.
Таблица 1.
Статистические тесты влияния функциональных признаков на эластичность жизненного цикла
SLA и лист N, две характеристики, которые представляют экономику быстрого листа, также были связаны с быстрым жизненным циклом.В регрессиях Дирихле обе эти характеристики были положительно связаны с эластичностью к плодовитости. Когда мы учли филогенетические отношения с регрессией PGLS, связь с эластичностью плодовитости оставалась значимой для N листа, но не для SLA. Эти черты были связаны с эластичностью индивидуального роста только тогда, когда мы добавляли формы роста или ковариаты биома. Мы обнаружили незначительно значимую отрицательную связь между SLA (но не N листа) и эластичностью к выживаемости ( P = 0.056), но эта связь не была значимой, когда мы добавили ковариаты формы роста и биома.
Взаимосвязи между функциональными характеристиками и исходными показателями жизнеспособности часто, но не всегда, соответствовали отношениям между характеристиками и эластичностями показателей жизнедеятельности. PGLS-регрессия исходных показателей жизнеспособности по функциональным признакам показала, что в некоторых случаях отношения признак-жизненный показатель соответствовали отношениям признак-эластичность (рис. 2, таблица 2, рис. S3 и таблицы S6 и S7).Например, увеличение массы семян, плотности древесины и продолжительности жизни листьев привело к значительному увеличению выживаемости и устойчивости к выживанию. Однако в других случаях отношения признак-жизненный цикл не соответствовали отношениям признак-эластичность: SLA и N листа не были связаны с коэффициентами плодовитости, но в значительной степени связаны с эластичностями по плодовитости. В целом, когда отношения функциональных признаков и показателей жизнеспособности были значимыми, соответствующие отношения признака и эластичности также были значимыми, но сильные связи функциональных признаков и показателей эластичности имели место, когда соответствующие отношения функциональных признаков и показателей жизнедеятельности были слабыми.
Рис. 2.
Сравнение влияния функциональных признаков на жизненно важные показатели и эластичность жизненного ритма. Стандартизованная величина эффекта — это значение Z (регрессия Дирихле) или t (регрессия PGLS) для следующих функциональных признаков: ( A ) масса семян, ( B ) плотность древесины, ( C ) лист. продолжительность жизни, ( D ) SLA и ( E ) лист N. Показатели жизнеспособности — это выживаемость (S), рост (G) и плодовитость (F). Отношения признак-эластичность были подобраны с использованием регрессии как Дирихле, так и PGLS, тогда как отношения признак-жизненный показатель были подобраны с использованием только регрессии PGLS.Пунктирными линиями показаны статистически значимые эффекты при α = 0,05.
Таблица 2.
Статистические тесты влияния функциональных признаков на показатели жизнеспособности
Обсуждение
Наши результаты показывают, что глобальные компромиссы в функциональных характеристиках растений предсказуемым образом приводят к компромиссам на основе жизненного цикла. Виды, вкладывающие средства в несколько крупных семян, имеют стратегии жизненного цикла, которые характеризуются более высоким влиянием на выживание и меньшим влиянием индивидуального роста и плодовитости на темпы роста популяции по сравнению с видами, производящими мелкие семена.Виды с более быстрой экономикой листьев и древесины также имеют тенденцию к более быстрой жизни, с более низкой эластичностью темпа роста популяции в отношении выживания и более высокой эластичностью в отношении индивидуального роста и плодовитости. Хотя функциональные черты объясняют лишь небольшую часть наблюдаемой вариабельности показателей жизнедеятельности и эластичности, наша способность обнаруживать любые существенные взаимосвязи замечательна, учитывая источники вариации, присущие нашему сравнительному подходу. Используя средние значения признаков для каждого вида, обычно измеряемые в местах, удаленных от места демографических наблюдений, мы игнорировали межпопуляционные различия как по функциональным признакам (37, 38), так и по демографии (39, 40).Тот факт, что наш грубый подход успешно выявил связи между функциональными чертами и стратегиями жизненного цикла, указывает на силу лежащих в основе причинных механизмов.
Остается без ответа важный вопрос об этих механизмах. Каким образом функциональные особенности, непосредственно влияющие только на ограниченный набор физиологических процессов и демографических показателей, могут объяснить вариации в общей истории жизни? Одно из возможных объяснений заключается в том, что затронутые процессы (например, производство семян и производительность всходов в случае массы семян) (32) особенно важны для их пригодности.Однако эмпирические данные указывают в противоположном направлении. Анализ эластичности обычно показывает, что для долгоживущих многолетних видов выживаемость взрослых особей оказывает гораздо большее влияние на приспособленность, чем плодовитость или выживаемость неполовозрелых особей (41). Более вероятное объяснение состоит в том, что изученные нами функциональные черты и процессы, на которые они влияют, развиваются совместно со многими другими чертами и процессами, которые в совокупности определяют историю жизни. Масса семян может объяснить большее разнообразие жизненного цикла, чем SLA (рис.2) не потому, что процессы, на которые он влияет, более важны, чем процессы, на которые влияет SLA, а просто потому, что масса семян является лучшим феноменологическим индикатором стратегии, скоординированной по функциям и стадиям жизни.
SLA может быть относительно плохим индикатором стратегии жизненного цикла из-за своей пластичности. Зависимость отношений SLA – эластичность от филогенетических корреляций или ковариат формы роста и биома согласуется с предыдущими исследованиями, показывающими, что SLA объясняет меньшие различия в демографических показателях, чем плотность древесины или размер семян (18, 23), и что значения SLA изменчивы и зависят от контекста. зависимые (42, 43).Мы также обнаружили, что SLA и лист N имеют более сильную связь с эластичностью к плодовитости, чем с ростом, что является неожиданным результатом, учитывая сильную положительную корреляцию между этими характеристиками и относительными темпами роста (1). Чувствительность эластичности плодовитости к этим признакам может указывать на то, что повышенные относительные темпы роста, которые сокращают время репродуктивной зрелости, имеют важные последствия для воспроизводства, и это иллюстрирует нашу гипотезу о том, что функциональные признаки влияют на жизненный цикл через сложный набор скоординированных процессов.
Значительного улучшения прогнозирующей способности взаимосвязей функциональная характеристика – эластичность можно достичь просто путем измерения функциональных характеристик и показателей жизнедеятельности в одно и то же время и в одном и том же месте. Еще один способ уменьшить необъяснимые вариации — сосредоточить внимание на видах в рамках определенных форм роста или в разных биомах. Мы включили эти ковариаты в наши модели, чтобы показать, что они не управляли взаимосвязями функциональных признаков и демографий, но во многих случаях они имели значительные независимые эффекты (таблицы S2 – S7) и часто улучшали нашу способность обнаруживать эффекты функциональных признаков.Например, добавление биома в качестве ковариаты к регрессии Дирихле изменило отношения между эластичностью роста и массой семян, SLA и N листа с незначительных на значимые. Наконец, будущие исследования могут объяснить большее разнообразие показателей жизнеспособности и эластичности, основывая прогнозы на нескольких функциональных характеристиках. Хотя мы не планировали наше исследование с учетом подхода множественной регрессии, у нас было достаточно видов с наблюдениями как за массой семян, так и за SLA, чтобы можно было провести первоначальное исследование.Простая линейная регрессия (логит-преобразованная) эластичности выживаемости по массе семян имела R 2 0,19, тогда как множественная регрессия эластичности выживаемости по массе семян и SLA увеличила R 2 до 0,25.
Наша вторая цель состояла в том, чтобы оценить использование необработанных показателей естественного движения населения для вывода о влиянии черт характера на историю жизни. Экологи в области народонаселения утверждали, что, поскольку приспособленность представляет собой чистый результат всех показателей жизнедеятельности, выводы о стратегиях жизненного цикла не могут быть основаны на полевых наблюдениях какой-либо индивидуальной жизненной скорости (44).Мы нашли частичную поддержку этой точки зрения. С одной стороны, значимость отношений между функциональными признаками и исходными показателями выживаемости всегда соответствовала отношениям между признаками и эластичностями выживаемости. С другой стороны, взаимосвязь между показателями сырой плодовитости и SLA и N листа не была значимой, тогда как взаимосвязь между этими характеристиками и эластичностью плодовитости была значимой. Для этих быстрорастущих экономических черт использование только необработанных данных об уровне естественного движения населения привело бы к недооценке влияния функциональных характеристик на жизненный цикл.
Объединив функциональные черты и модели населения, мы смогли ответить на давние вопросы о влиянии функциональных черт на приспособленность и историю жизни, используя количественные методы, разработанные в сравнительной демографии. Мы обнаружили, что ранее задокументированные взаимосвязи между чертой и показателем жизнеспособности распространяются на эластичность показателей жизнедеятельности, напрямую связывая черты со всесторонней оценкой индивидуальной приспособленности и подтверждая центральное предположение об экологии на основе черт. Эмпирические доказательства сильной связи между простыми анатомическими и морфологическими признаками и стратегиями жизненного цикла ставят основанные на признаках подходы на прочную основу и должны способствовать применению функциональных признаков в популяционной экологии.В конечном счете, подход функциональных признаков может помочь нам определить анатомические и физиологические основы эволюции жизненного цикла, важную нерешенную проблему на стыке экофизиологии, популяционной экологии и эволюционной биологии.
Материалы и методы
Функциональные признаки растений.
Мы получили данные о пяти обычно измеряемых функциональных признаках из TRY, глобального хранилища данных о признаках растений (35): масса семян (миллиграммы), плотность древесины (миллиграммы на миллиметр 3 ), продолжительность жизни листьев (месяцы), SLA ( миллиметры 2 на миллиграмм) и листовой N (миллиграммы на грамм).Мы извлекли данные о функциональных признаках для любых видов многолетних растений в TRY, по которым у нас были демографические данные (см. Ниже). После удаления повторяющихся наблюдений за функциональными признаками и выбросов (значения, превышающие 3 SD от среднего значения признака для каждого вида), мы рассчитали средние значения для каждого функционального признака, наблюдаемого для каждого вида.
Показатели жизнедеятельности и эластичность.
Мы получили демографическую информацию из базы данных COMPADRE Plant Matrix (45) (www.compadre-db.org/), которая содержит матрицы прогнозов для более чем 500 видов растений, а также таксономические и экологические ковариаты.Эти матрицы прогнозов суммируют основные демографические процессы (т. Е. Показатели жизнедеятельности), которые определяют скорость роста популяции (λ), такие как выживаемость, изменения стадии / размера, семенной банк и вегетативный покой, клональное размножение и половое размножение (27).
Для каждого из 222 видов, которые совпадали с базой данных TRY, мы оценили матрицу средних значений по элементам для всех лет и популяций, которые наблюдались в контрольных условиях (мы исключили экспериментальные обработки).Если для данного вида было доступно более одной матричной модели, мы выбирали модель с наибольшей пространственной и временной репликацией. В случае, если две модели для одного и того же вида имели равную пространственную и временную репликацию, мы выбрали модель с наибольшим размером матрицы, потому что модели с более высокой размерностью матрицы предлагают описание динамики популяции с более высоким разрешением (46). Мы проигнорировали опубликованные исследования, в которых матричная модель не включала меры плодовитости, поскольку для расчета эластичности требуется информация обо всем жизненном цикле (27).
Мы разложили показатели жизнеспособности для каждого вида на выживаемость, плодовитость и рост, и мы рассчитали эластичность λ по отношению к этим показателям приживаемости, используя уравнения 7–11 в исх. 30. Эта декомпозиция учитывает, что вероятности перехода, содержащиеся в отдельных элементах матрицы, часто представляют более одного показателя жизнедеятельности. Например, недиагональные записи обычно представляют вероятность выживания и прогрессирования или регресса. Показатели естественного движения населения дают представление о реальной динамике населения (т.е., коэффициенты выживаемости, роста и воспроизводства), тогда как эластичности обеспечивают относительное влияние этих демографических процессов на темпы роста населения (47).
На первый взгляд, измерение роста с использованием матричных моделей популяции качественно отличается от измерений относительной скорости роста, традиционно используемых в исследованиях функциональных признаков, таких как биомасса (граммы граммы -1 год -1 ) или диаметр стержня (сантиметры сантиметры). -1 год -1 ).Наша мера роста представляет собой среднюю вероятность того, что человек перейдет из одного класса размера или стадии в другой за единичный интервал времени, который в каждом случае равнялся 1 году. В нашем наборе данных эти классы размера и стадии определяются множеством способов, таких как количество листьев, диаметр стебля, диаметр розетки, высота по возрасту и / или стадия развития. Независимо от того, как определены эти размеры и стадии, единицы роста идентичны и представляют собой относительную скорость роста (индивидуумы -1 год -1 ).Фактически, независимо от того, определяются ли относительные темпы роста в терминах биомассы, диаметра стебля или вероятностей перехода, единицы измерения — это просто год -1 . Важным преимуществом количественной оценки роста с помощью матричных моделей является то, что этот подход учитывает изменение относительных темпов роста в зависимости от размера или стадии. Хотя решения о количестве классов размера / стадии для включения могут влиять на исходные темпы роста (меньшее количество классов означает более высокую вероятность перехода), общая эластичность к росту не зависит от размерности матрицы и обеспечивает безразмерный индекс влияния роста на приспособленность. .
Поскольку нашей демографической единицей для сравнения был генет (генетически идентичный индивид), мы суммировали клональное размножение с индивидуальным прогрессированием / ростом, регрессом / сокращением и вегетативным покоем. Изменения размера влияют на чувствительность и эластичность по отношению к репродуктивной ценности участвующих размерных классов (например, рост с класса x до x + 1 или снижение до x — 1). Положительный и отрицательный рост имеют одинаковый чистый эффект на репродуктивную ценность человека размером x (27), но знак эффекта зависит от направления изменения (симметрия уравнений 8 и 9 в исх.30). Мы решили количественно оценить влияние изменений в размере на рост населения независимо от их знака, а не позволять одному направлению не учитывать другое, что может создать необоснованную возможность изменения размера, не влияющего на темпы роста населения. Прорастание семян до рассады в течение годового периода производства семян и взносы банка семян были включены в плодовитость.
Хотя чувствительность к показателю естественного движения населения полезна для оценки вклада в темп роста населения наблюдаемых (т.е., положительные) и гипотетически положительные матричные коэффициенты (т. е. ненаблюдаемые переходы равны нулю) (48, 49), мы не представляем результаты для чувствительности показателей естественного движения населения, поскольку их неаддитивность не позволяет проводить четкие сравнения между различными показателями естественного движения населения. Кроме того, биологическое значение гипотетических переходов трудно оценить, имея дело с многочисленными переходами для сотен видов.
Мы не включали размер матрицы в качестве ковариаты, потому что, хотя известно, что он влияет на суммарную эластичность элементов матрицы (46, 50), он оказывает минимальное влияние на суммарную эластичность показателей естественного движения населения (51), что дает нам уверенность. что наши оценки надежны.Кроме того, различия между видами в размерности матрицы часто отражают реальные различия в истории жизни. Например, матрицы, используемые для моделирования деревьев, часто больше, чем матрицы, используемые для кустарников или трав.
Филогения.
Мы попытались получить филогению для 222 видов в нашем наборе данных, используя два метода: генератор деревьев на основе таксономии Национального центра биотехнологической информации в iTOL (http://itol.embl.de/index.shtml) и phyloGenerator (52 ). Однако GenBank не содержит информации по большинству видов в нашем наборе данных, и даже если в GenBank можно было найти близкородственные виды и использовать их в качестве временных суррогатов, полученное дерево оказалось трудно разрешить с помощью предложений phyloGenerator.Следовательно, мы прибегли к составлению списка видов с их признанными семействами в PHYLOMATIC v2 (53) (прекращено 10 января 2013 г .; Phylomatic v3 теперь находится на http://phylodiversity.net/phylomatic/). Затем получившееся дерево было вручную преобразовано в MESQUITE (http://mesquiteproject.org) с использованием информации с веб-сайта Angiosperm Phylogeny Website (www.mobot.org/MOBOT/research/APweb/) и конкретных подробных исследований филогенетических взаимоотношений внутри семей, о которых идет речь. на веб-сайте Angiosperm Phylogeny или найдены нами самостоятельно.Поскольку не все роды в нашем наборе данных были включены в филогенетические исследования (у многих видов отсутствует информация о последовательности ДНК в GenBank), мы также полагались на признанные таксономические различия, основанные на морфологических и фенотипических признаках. Хотя такое использование традиционной таксономии может показаться произвольным, различия филогенетических расстояний невелики и не меняют наблюдаемые взаимосвязи, что мы подтвердили, проведя анализ на разных этапах построения окончательного филогенетического дерева.Филогенетические расстояния максимально разрешенного филогенетического дерева были интерполированы с использованием функции bladj программы PHYLOCOM (54) с оценочными возрастами узлов, приведенными в ссылке. 55.
Анализ.
Мы использовали регрессии Дирихле (56, 57), чтобы учесть композиционный характер эластичностей жизненного цикла (для каждого вида эластичности по выживаемости, росту и плодовитости суммируются с единицей), и мы использовали регрессию PGLS для контроля отсутствия независимости видов с общим происхождением.Мы обнаружили, что регрессия PGLS, которая игнорирует композиционную природу эластичности, часто демонстрирует значительные взаимосвязи между характеристикой и эластичностью, тогда как регрессия Дирихле — нет. Например, сильная взаимосвязь между плотностью древесины и эластичностью выживания неизбежно приводит к взаимосвязи с индивидуальным ростом или эластичностью плодовитости, которые кажутся значимыми при независимом рассмотрении. Поэтому мы последовательно интерпретировали результаты регрессий Дирихле и PGLS. Если регрессия Дирихле указала на значительную взаимосвязь признак-эластичность, мы исследовали PGLS, чтобы убедиться, что этот эффект не был просто результатом филогенетических корреляций.Мы подбираем второй и третий наборы моделей, в которые добавили форму роста или биом в качестве категориальных независимых переменных. Для плотности древесины регрессия Дирихле, которая включала биом, была чрезмерно параметризована (биом пустыни был представлен только одним видом). Поэтому мы подбираем модель плотности древесины плюс биома после удаления синглтона пустыни. Чтобы регрессировать показатели жизнеспособности по функциональным признакам, мы использовали PGLS, как описано выше, после логит-преобразования коэффициентов выживаемости и логарифмических коэффициентов роста и плодовитости.
Мы реализовали регрессию Дирихле в R v.2.15.1 (58) с помощью пакета DirichletReg. Мы реализовали регрессию PGLS в R с помощью пакета ape (59). Мы использовали модель эволюции броуновского движения, а не модель Орнштейна – Уленбека, потому что в очень немногих случаях, когда результаты различались, модель броуновского движения была более консервативной в отношении эффектов признаков. Для регрессий PGLS мы независимо рассматривали эластичность по выживаемости, росту и плодовитости и преобразовывали их с помощью логита.
Благодарности
Мы благодарим Люка Захмана и Энди Кляйнхесселинка за помощь в сопоставлении данных о признаках, а также Дэвида Кунса, Натана Крафт, Джонатана Левина и Дженнифер Уильямс за полезные комментарии к более ранней версии рукописи. P.B.A. был поддержан грантом Национального научного фонда DEB-1054040. Р.С.-Г. благодарит за поддержку Институт демографических исследований Макса Планка. A.C. был поддержан Государственным университетом Юты. J.S.H. был поддержан стипендией выпускников Национального научного фонда.C.M.-A. и М.Ф. были поддержаны Плимутским университетом.
Сноски
Авторы: P.B.A., R.S.-G., A.C., J.S.H. и J.R.-M. спланированное исследование; P.B.A., R.S.-G., A.C., J.S.H., J.R.-M., C.M.-A. и M.F. проведенное исследование; П. Б. А., Р. С.-Г. и М.Ф. проанализированные данные; и P.B.A., R.S.-G., J.S.H., J.R.-M. и M.F. написал газету.
Авторы заявляют об отсутствии конфликта интересов.
Эта статья представляет собой прямое представление PNAS.
Размещение данных: данные размещены в Data Dryad, http://doi.org/10.5061/dryad.8g252.
Эта статья содержит вспомогательную информацию на сайте www.pnas.org/lookup/suppl/doi:10.1073/pnas.1315179111/-/DCSupplemental.
Яркая история комнатных растений — Советы по уходу за растениями и многое другое · La Résidence
Согласно Британской энциклопедии, практика домашнего садоводства с использованием горшечных растений восходит к древним грекам и римлянам.Более старые цивилизации, такие как Древний Египет, Индия и Китай, также использовали растения в горшках, но в основном на открытом воздухе и во дворах. В горшках преобладала терракота — за исключением древних римлян, которые тяготели к мраморным горшкам (так шикарно!).
В японской, вьетнамской и китайской культурах существуют уникальные традиции создания карликовых деревьев для декоративных целей, известных под разными названиями: Хон Нон Бо, Пенцзин и Бонсай. Эти растения были спроектированы так, чтобы отражать форму взрослых деревьев в природе, и часто сопровождались декоративными камнями и даже водными элементами для имитации природных ландшафтов.
В Висячих садах Вавилона на территории современного Ирака даже был особый сад, специально созданный для жены царя Навуходоносора II. Он продемонстрировал листву ее родины с финиковыми деревьями, статными кедрами и лугами.
Эпоха Возрождения и за его пределами
Средневековье не было подходящим местом для выращивания комнатных растений или, можно утверждать, для чего угодно! В тот период в Европе культивирование комнатных растений в основном ограничивалось монастырями, а растения выращивались для практических целей (в основном для еды и медицины), а не для удовольствия или эстетики.
Итак, слава богу, за Ренессанс и культурные преобразования, которые он возвестил! В эпоху Возрождения в моду вошли оранжерей , а глобальные исследования а-ля Христофора Колумба означали, что приток листвы из нового мира был принесен домой, чтобы продемонстрировать их в Европе.
Лучшие садоводы: 1840–1930
С индустриализацией и викторианской эпохой дома обычных людей стали больше напоминать дома, которые мы знаем сегодня.Они были теплее благодаря центральному отоплению от угольных или железных печей, что позволило людям поднять выращивание комнатных растений на новый уровень. Папоротники были исключительно популярны из-за своей густой роскошной листвы, и их часто помещали в контейнеры в виде колонн, которые назывались jardiniere. Другие растения, которые мы до сих пор знаем и любим (и продаем на Léon & George!), Также пользовались популярностью, например, Cast Iron Plants и Kentia Palms.
Границы | От редакции: Особенности жизненного цикла патогенов растений и адаптация к ограничениям окружающей среды
История жизни является ключевым понятием в эволюционной биологии и экологии.Это соответствует повествованию о различных событиях, перемежающих существование организма от его рождения до его смерти (Begon et al., 2006; Michalakis et al., 2016). На протяжении всей жизни живые организмы приобретают ресурсы, которые они активно находят или извлекают из окружающей среды, а затем распределяют их для различных функций: развития, выживания, распространения и воспроизводства (Roff, 1992; Stearns, 1992; Michalakis, 2009). Черты, влияющие на время и амплитуду этих дилемм распределения, определяются как черты жизненного цикла.Черты жизненного цикла часто являются определяющими для людей (Kingsolver and Pfennig, 2007). Они влияют на пространственную и временную динамику заболевания и, следовательно, на генетическое разнообразие и структуру популяций патогенов (Barrett et al., 2008; Michalakis et al., 2016). Эти детерминанты участвуют в способности патогенов адаптироваться к различным экологическим факторам, включая изменения биотического и абиотического воздействия, а также прямое или косвенное взаимодействие с другими штаммами или видами патогенов, совместно встречающимися на одном и том же хозяине (Michalakis et al., 2016; Tollenaere et al., 2016). Для патогенов растений эти признаки можно разделить на две категории: те, которые участвуют в фазе эпидемии, на хозяине или внутри хозяина, и те, которые связаны с фазой выживания, часто вне хозяина. Между этими двумя фазами может происходить компромисс, что приводит к последствиям для динамики, эволюции и видообразования эпидемии (Pariaud et al., 2009; Hamelin et al., 2011). Понимание процессов, поддерживающих вариабельность жизненных характеристик патогенов растений, является центральным вопросом эволюционной экологии и серьезной проблемой при разработке стратегий борьбы с болезнями (Galvani, 2003; Grenfell et al., 2004).
Целью данной темы исследования был сбор исследовательских работ с обширным взглядом на особенности жизненного цикла патогенных микроорганизмов растений и адаптацию к ограничениям окружающей среды. Были рассмотрены особенности жизненного цикла как фазы эпидемии, так и фазы выживания, и проверенные экологические ограничения включали абиотические (температура, осадки, водный стресс) и биотические (среда обитания, сорт, зрелость листьев, коинфекция) факторы. Среди 10 принятых статей были рассмотрены три типа патогенов растений: грибы (семь статей), нематоды (две статьи) и оомицеты (одна статья).
Что касается фазы эпидемии, четыре основные темы были охвачены исследовательскими работами, принятыми в этом специальном выпуске: i) адаптация хозяина, ii) влияние физиологического статуса хозяина, iii) адаптация к климатическим условиям и iv) эффектов коинфекции.
Как адаптация хозяина влияет на эволюцию жизненных черт, в частности, анализировалось путем изучения влияния устойчивых сортов на несколько жизненных черт. В аскомицете Colletotrichum gloeosporioides изучалось влияние количественной устойчивости хозяина на эволюцию патогена путем оценки разнообразия популяций в полевом масштабе как по нейтральным маркерам, так и по патогенным признакам (Frezal et al.). Генетическая структура популяции выявила значительное влияние клонального воспроизводства в эволюции C. gloeosporioides и низкую скорость миграции между полями. Результаты тестов перекрестной инокуляции показали, что агрессивность грибковых клонов, по-видимому, развивалась за счет накопления компонентов, специфичных для каждого сорта водяного ямса, что свидетельствует об адаптации к их сорту-хозяину. Несмотря на оставшиеся признаки адаптации к бывшему широко культивируемому хозяину, адаптация к текущим сортам была четко изображена.Подобный паттерн локальной адаптации к доминирующему сорту ранее был отмечен у возбудителя фитофтороза картофеля Phytophthora infestans (Andrivon et al., 2007). В своем исследовании Mariette et al. исследовали, имеет ли место теперь классическая отрицательная связь между размером и численностью потомства у этого оомицета, P. infestans , и влияет ли на компромисс сорт картофеля или хозяин, являющийся источником патогена (томат и картофель). Они подтвердили существование компромисса и показали, что на него не повлиял ни один из этих биотических факторов.Наблюдаемый полифенизм по этим признакам в популяциях P. infestans будет способствовать сосуществованию различных репродуктивных стратегий.
В последние годы динамика внутри хозяина была определена как главный компонент эпидемиологической и эволюционной динамики патогенов (Alizon et al., 2009). Известно, что у растений характеристики тканей хозяина сильно зависят от возраста ткани. В частности, в зависимости от возраста тканей доступность питательных веществ, а также уровни защиты могут значительно различаться, что оказывает противоположное влияние на успех инфекции (Kus et al., 2002; Аль-Наими и др., 2005). Онтогенная устойчивость к мучнистой росе ( Erysiphe necator) на Vitis vinifera была изучена Calonnec et al. Они показали, что были затронуты три изученных патогенных признака (эффективность заражения, споруляция и рост мицелия), и их вариабельность сильно коррелировала с возрастом листьев. Споруляция более тесно коррелировала с вариациями содержания сахара, а эффективность заражения — с содержанием воды в листьях, что позволяет предположить, что онтогенная резистентность листьев виноградной лозы, по-видимому, является неизменным физиологическим процессом, чем E.necator способен обойти, ограничивая свое развитие тонкой тканью. Аналогичным образом Maupetit et al. проверили, как доступность питательных веществ и уровень защиты влияют на развитие Melampsora larici-populina (эффективность заражения, латентный период, размер урединии, количество мицелия, скорость споруляции, способность споруляции и объем спор). Они показали, что M. larici-populina была более агрессивной на более зрелых листьях, на что указывают более широкие урединии и более высокая скорость споруляции.Напротив, содержание фенолов (флавонолы, эфиры гидроксикоричной кислоты и салициноиды) отрицательно коррелировало с размером урединии и скоростью споруляции, что свидетельствует о том, что приспособленность патогена, по-видимому, в большей степени ограничивается конститутивным уровнем защиты растений, чем доступностью питательных веществ, что видно по уменьшению в споруляции.
Подобно другим организмам, патогенам необходим определенный диапазон температуры и влажности, чтобы проникнуть и развиваться на своем растении-хозяине (Agrios, 2005).Эти два параметра, в частности, могут изменять прорастание спор, колонизацию хозяина, образование спор и распространение грибковых патогенов (Fitt et al., 2006). Десятилетия исследований позволили получить обширные знания и лучшее понимание сезонного воздействия температуры, осадков и влажности на болезни, поражающие основные продовольственные культуры (Garrett et al., 2011; Pautasso et al., 2011; Donatelli et al., 2017). В этом специальном выпуске были изучены два основных параметра: изменение температуры и влияние водного стресса.Что касается влияния температуры, Vaumourin и Laine разработали исследование Podosphaera plantaginis и показали, что температура оказывает значительное влияние на все измеренные характеристики жизненного цикла, предполагая, что влияние температуры на жизненные черты было как прямым, так и прямым. опосредовано взаимодействием между генотипом и температурой. Аналогичным образом Mariette et al. показал на P. infestans , что отрицательная взаимосвязь между размером и численностью потомства сохраняется для всех тестируемых температур.Другие исследования с использованием цистных нематод показали влияние температуры на различные жизненные черты. Действительно, Fournet et al. показали сильное влияние температуры на жизненные признаки нематоды свекловичной цисты Heterodera schachtti . Хотя на размножение нематод по-разному влияла температура, поскольку благоприятные условия для хозяина также благоприятны для паразита, влияние температуры на отрождение зависело от происхождения популяций, отделяющих южные от северных европейских популяций.Используя подход сканирования генома, Gendron St-Marseille et al. оценили адаптивный потенциал цистовой нематоды сои Heterodera glycines . Генотипирование путем секвенирования 64 популяций H. glycines позволило идентифицировать 15 локусов при отборе по климатическим или географическим переменным. Наконец, что касается воздействия водного стресса, для патосистемы Arabidopsis thaliana / Alternaria brassicicola был разработан транскриптомный и метаболомный подход для изучения уровня восприимчивости гриба к водному стрессу и его влияния на его способность передавать семена (N ‘ Guyen et al.). Эти подходы привели к идентификации специфических белков (гидролипина), участвующих в устойчивости к водному стрессу и передаче семян, но не в агрессивности патогена.
Способность патогена закрепляться и расти на своем хозяине может резко измениться при одновременном заражении другими патогенами (другими штаммами или видами патогенов, одновременно встречающимися на одном и том же хозяине). Распределение по анамнезу может измениться при коинфекции, влияя на эволюционный потенциал и эпидемиологическую динамику патогенов (Tollenaere et al., 2016; Лайне и Мякинен, 2018). В своем исследовании Ваумурин и Лайне обнаружили, что коинфекция только изменила количество производимых половых структур покоя, но не изменила производство бесполых форм. Исследования коинфекций, оценка грибов с общими характеристиками и видов-хозяев создают проблему для традиционной диагностики заболеваний и последующих стратегий лечения. Действительно, отсутствие мощных инструментов для одновременного различения и количественной оценки этих типов патогенов является серьезным ограничением.В своем исследовании Abdullah et al. разработали дуплексный анализ ПЦР в реальном времени для количественного определения сопутствующих патогенов пшеницы, Pyrenophora tritici-repentis и Parastagonospora nodorum. Полезность метода была продемонстрирована на полевых образцах сорта, чувствительного к обоим патогенам. В то время как визуальная и посевная диагностика предполагали присутствие только одного из видов патогенов, анализ выявил не только присутствие обоих сопутствующих патогенов (что позволило выявить бессимптомно), но также позволило количественно оценить относительную численность патогенов в зависимости от тяжести заболевания. .
Этап выживания за пределами хозяина часто упускается из виду из-за сложности его изучения. Это ключевой этап жизненного цикла патогена. Все патогены зависят от жизненного цикла, где, несмотря на существование неактивных форм, могут происходить эволюционные процессы. Таким образом, в зависимости от статуса хозяина (многолетнее или однолетнее растение-хозяин) патогены должны разработать широкий спектр стратегий для преодоления чередования присутствия / отсутствия (и / или модификации физиологической активности) своего хозяина (Agrios, 2005). ).Следовательно, эта фаза выживания важна, поскольку она может повлиять на размер популяции патогена и привести к возникновению узких мест. В этом специальном выпуске исследование, разработанное Bardin et al. исследовали повсеместность широкого круга хозяев некротрофного гриба Botrytis cinerea за пределами сельскохозяйственных угодий и определили, можно ли фенотипически и филогенетически отличить популяции в этих естественных местообитаниях от популяций, выделенных из больных сельскохозяйственных культур.Их результаты показали, что штаммов B. cinerea , отобранные на различных несельскохозяйственных субстратах, были генетически и фенотипически подобны штаммам, отобранным на сельскохозяйственных субстратах. Эти результаты предполагают, что весьма разнообразные популяции этого патогена растений сохраняются за пределами сельского хозяйства в сочетании с субстратами, отличными от культурных растений, и что этот компонент их жизненного цикла совместим с его способностью сохранять свой потенциал в качестве патогена растений. Фаза выживания также рассматривалась Vaumourin и Laine, поскольку коинфекция влияет на количество образованных половых структур покоя, а в Fournet et al., поскольку они также учитывали вылупление после хранения при различных температурах, моделируя условия выживания в период межкультурного посева.
Рассмотрение признаков патогенов растений в контексте эволюции признаков жизненного цикла является многообещающим направлением исследований, которое все еще находится в зачаточном состоянии. Мы надеемся, что эта тема исследования будет способствовать развитию этого направления исследований и станет ценным ресурсом для исследователей, работающих в области эволюционной экологии популяций патогенов растений.
Вклад авторов
CL, JM, CM, OF и VR внесли существенный непосредственный вклад в работу и одобрили ее для публикации.
Конфликт интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы благодарим всех авторов, вовлеченных в эту тему исследования, за их отличный и разнообразный вклад.
Ссылки
Agrios, G. N. (2005). Патология растений — 5-е изд. (США: Elsevier Academic Press).2005.
Google Scholar
Ализон, С., Херфорд, А., Мидео, Н., ван Баален, М. (2009). Эволюция вирулентности и гипотеза компромисса: история, текущее положение дел и будущее. J. Evol. Биол. 22, 245–259. doi: 10.1111 / j.1420-9101.2008.01658.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Аль-Наими, Ф. А., Гаррет, К. А., Бокус, В. В. (2005). Конкуренция, содействие и дифференциация ниш у двух листовых патогенов. Oecologia 143, 449–457. doi: 10.1007 / s00442-004-1814-x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Andrivon, D., Pilet, F., Montarry, J., Hafidi, M., Corbière, R., Achbani, E.H., et al. (2007). Адаптация Phytophthora infestans к частичной устойчивости картофеля: данные французского и марокканского населения. Phytopathol 97, 338–343. doi: 10.1094 / PHYTO-97-3-0338
CrossRef Полный текст | Google Scholar
Barrett, L.Г., Тралл, П. Х., Бердон, Дж. Дж., Линде, К. С. (2008). История жизни определяет генетическую структуру и эволюционный потенциал взаимодействий паразит-хозяин. Trends Ecol. Evol. 23, 678–685. doi: 10.1016 / j.tree.2008.06.017
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бегон, М., Таунсенд, К. Р., Харпер, Дж. Л. (2006). Экология: от индивидов к экосистемам (Мальден, Массачусетс: Blackwell Pub.).
Google Scholar
Донателли, М., Магарей, Р. Д., Брегальо, С., Виллоккет, Л., Виш, Дж. П. М., Савари, С. (2017). Моделирование воздействия вредителей и болезней на сельскохозяйственные системы. Agric. Syst. 155, 213–224. doi: 10.1016 / j.agsy.2017.01.019
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фитт, Б. Д. Л., Хуанг, Ю.-Дж., ван ден, Бош Ф., Дж. С., Вест (2006). Сосуществование родственных видов патогенов на пахотных культурах в пространстве и времени. Ann. Rev. Phytopathol. 44, 163–182. DOI: 10.1146 / annurev.phyto.44.070505.143417
CrossRef Полный текст | Google Scholar
Гальвани, А. П. (2003). Эпидемиология встречается с эволюционной экологией. Trends Ecol. Evol. 18 (3), 132–139.
Google Scholar
Гаррет, К. А., Нита, М., Де Вольф, Э. Д., Эскер, П. Д., Гомес-Монтано, Л., Спаркс, А. Х. (2011). «Патогены растений как индикаторы изменения климата» в издании «Изменение климата» (2-е изд.): Наблюдаемые воздействия на планету Земля, . Эд. Летчер, Тревор М.(Амстердам, Нидерланды: Elsevier BV), 325–338.
Google Scholar
Гренфелл, Б. Т., Пибус, О. Г., Гог, Дж. Р., Вуд, Дж. Л. Н., Дейли, Дж. Н., Мамфорд, Дж. А., Холмс, Е. К. (2004). Объединение эпидемиологической и эволюционной динамики патогенов. Наука 303 (5656), 327–332.
PubMed Аннотация | Google Scholar
Hamelin, F., Castel, M., Poggi, S., Andrivon, D., Mailleret, L. (2011). Сезонность и эволюционная дивергенция паразитов растений. Экология 92 (12), 2159–2166. doi: 10.1890 / 10-2442.1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Kingsolver, J. G., Pfennig, D. W. (2007). Закономерности и сила фенотипического отбора в природе. BioScience 57 (7), 561–572. doi: 10.1641 / B570706
CrossRef Полный текст | Google Scholar
Кус, Дж. В., Затон, К., Саркар, Р., Камерон, Р. К. (2002). Возрастная устойчивость Arabidopsis является регулируемым развитием ответом Ddfense на Pseudomonas syringae . Растительная клетка 14, 479–490. doi: 10.1105 / tpc.010481
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Michalakis, Y., Charmantier, A., Gaillard, J.-M., Sorci, G., Tully, T., Ronce, O. (2016). «Evolution des traits d’histoire de vie», в Biologie évolutive (Louvain-la-Neuve: De Boeck Supérieur), 373–422.
Google Scholar
Михалакис Ю. (2009). «Паразитизм и эволюция особенностей жизненного цикла», в Экология и эволюция паразитизма (Oxford: Oxford University Press), 19–30.
Google Scholar
Парио, Б., Равин, В., Халкет, Ф., Гойо, Х., Карлье, Дж., Ланну, К. (2009). Агрессивность и ее роль в адаптации патогенов растений. Plant Pathol. 58, 409–424. doi: 10.1111 / j.1365-3059.2009.02039.x
CrossRef Полный текст | Google Scholar
Паутассо, М., Доринг, Т. Ф., Гарбелотто, М., Пеллис, Л., Джегер, М. Дж. (2011). Воздействие изменения климата на болезни растений — мнения и тенденции. Eur. J. Plant Pathol. 133, 295–313. doi: 10.1007 / s10658-012-9936-1
CrossRef Полный текст | Google Scholar
Рофф, Д. А. (1992). Эволюция историй жизни: теория и анализ (Нью-Йорк: Чепмен и Холл).
Google Scholar
Стернс, С. К. (1992). Эволюция историй жизни (Oxford: Oxford University Press).
Google Scholar
Tollenaere, C., Susi, H., Laine, A.-L. (2016). Эволюционные и эпидемиологические последствия множественной инфекции растений. Trends Plant Sci. 21, 80–90. doi: 10.1016 / j.tplants.2015.10.014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Музей Генри Б. Планта — История
В 1880-х годах завод Генри Брэдли строил империю железных дорог, пароходов и отелей. Он хотел, чтобы в этой империи был дворец, и этим дворцом был отель «Тампа-Бэй». Отель был построен Плант лично, а не инвесторами, по цене 2 500 000 долларов и еще 500 000 долларов на меблировку.На строительство потребовалось два года, площадь 6 акров и длина 1/4 мили. Его 511 номеров были одними из первых во Флориде, полностью оснащенных электричеством. Рекламируемое как полностью огнестойкое, здание содержало заливной бетон, армированный рельсами и кабелями между этажами. В здании были все самые современные предметы роскоши, включая бильярдную, парикмахерскую, услуги по чистке обуви, салон красоты, цветочный магазин, телеграф, столовую для торжественных мероприятий, Большой салон, Музыкальный зал с оркестром и телефоны во всех номерах.
Когда Генри Б. Плант привел железную дорогу в Тампу в 1884 году, он открыл дверь для процветания многих предприятий. Железная дорога была центром экономики города. Из центра он направлялся в Порт-Тампа-Сити, откуда можно было сесть на пароход Plant Line до Мобила, Алабамы, Ямайки, Кубы или Бермудских островов. Инвесторы с подозрением относились к строительству отеля такого масштаба, как задумал Завод, поэтому Плант решил, что построит его сам. Строительство отеля Tampa Bay обошлось примерно в 2 500 000 долларов, а на отделку — 500 000 долларов.Рекламируемый как абсолютно пожаробезопасный, благодаря стальным рельсам для армирования и бетонной конструкции, он состоял из 511 комнат. Некоторые помещения представляли собой апартаменты от трех до семи комнат, которые обеспечивали уровень роскоши, невиданной ранее в Тампе. В большинстве комнат были отдельные ванные комнаты, во всех было электричество и телефоны. Гости и багаж перемещались между пятью этажами на двух первых пассажирских и грузовых лифтах Флориды. Гости наслаждались атмосферой пьянящей роскоши, окруженной экзотической мебелью, фарфором, венецианскими зеркалами и скульптурами, подобранными в Европе г-ном.и миссис Плант. Декор представлял собой коллекцию, демонстрирующую такой изысканный вкус, что один писатель описал ее как — шкатулку с драгоценностями, в которую было собрано бесконечное количество драгоценных камней.
Открытый с декабря по апрель на протяжении 1890-х годов, отель был оживленным местом, где проводились великолепные балы, чаепития и организовывались охоты во время зимнего социального сезона. Гости наслаждались множеством развлечений, в том числе охотой на дичь, рыбной ловлей в пресной и соленой воде, парусным спортом, греблей и греблей на каноэ.В их распоряжении были велосипеды и экипажи. Рикши были доступны для экскурсий по собственности или дневной поездки, чтобы увидеть и быть увиденным. В отеле также есть поле для гольфа, корты для тенниса и шаффлборда, бильярд, крокет и даже ипподром. В течение недели в Music Room проходили грандиозные балы и оркестровые концерты. Казино Tampa Bay вмещало 2000 человек и выставляло счета национальным и международным исполнителям, таким как Нелли Мельба, Сара Бернар, Джон Филип Соуза и Анна Павлова. Казино также служило спа-центром с крытым бассейном с подогревом, расположенным под съемными половицами, чтобы создать оазис расслабления.Большой салон, или гостиная, был местом вдохновения, наполненным европейскими статуями и зеркалами в венецианском стиле. В 1892 году писатель The New York Times Уильям Дрисдейл заметил, что джентльменов пера, которые могут написать пару колонок о заснеженной горной вершине и впасть в экстаз на десять страниц после восхода солнца, должны стоять посреди Тампы. Bay Hotel и позвольте их безупречному английскому течению безудержно.
Широкая веранда на восточной стороне отеля предоставляет гостям спокойную обстановку, из которой открывается вид на обширные сады отеля.После формального ужина из восьми блюд с прекрасным вином и изысканной кухней в столовой гости могли прогуляться по серпантинным дорожкам, пролегающим через экзотический ландшафт тропической флоры и садовых скульптур.